An dieser Stelle finden Sie über den Inhalt des Buchs „Evolution - ein kritisches Lehrbuch“ hinaus einen zusätzlichen Text zum Kapitel 9.3 „Molekulare Grundlagen von Evolutionssprüngen“.
Vorbemerkung. Dieser Text stammt aus der Auflage von 2001. Er ist zwar unseres Erachten nach wie korrekt, es ist aber geplant, den Text durch neuere Literatrur zu aktualisieren. Für Hinweise sind wir dankbar. Versuche zur Entstehung von organischen Molekülen aus anorganischen Bestandteilen (Entstehung des Lebens in „Ursuppen“) werden in IV.7 besprochen. Hier soll von Experimenten die Rede sein, bei denen „Darwinsche Evolution“ im Reagenzglas beobachtet werden kann: Unter bestimmten Bedingungen sind Selektionsprozesse nicht an Zellen oder Organismen gebunden, sondern können auch in vitro in künstlichen Systemen ablaufen. Dabei benutzt man Proteine und Nukleinsäuren, die aus Bakterien isoliert wurden. Besonders gut lassen sich Spezialisierungen demonstrieren, bei denen unter bestimmten Selektionsbedingungen nicht gebrauchte Strukturen verloren gehen. Doch auch Änderungen vorhandener Strukturen werden beobachtet. Können dadurch Erkenntnisse gewonnen werden, welche man an ganzen Zellen nicht so leicht gewinnen kann? Welche Beobachtungen in solchen Systemen sprechen für die Entstehung ganz neuartiger Strukturen? Das Bakterienvirus Q
|
|
Wir werden zuerst ein Experimentalsystem kennenlernen, das auf der RNS-Replikase von Q![]() beruht. Q
beruht. Q![]() ist ein sehr kleines Bakterienvirus, das Escherichia coli-Zellen befällt. Es trägt als Erbsubstanz ein einzelsträngiges, kurzes +RNS-Molekül. Dieses enthält nur sehr wenige Gene, eines davon trägt die Information für die Q
ist ein sehr kleines Bakterienvirus, das Escherichia coli-Zellen befällt. Es trägt als Erbsubstanz ein einzelsträngiges, kurzes +RNS-Molekül. Dieses enthält nur sehr wenige Gene, eines davon trägt die Information für die Q![]() -Replikase. Dieses Enzym stellt vom +RNS-Strang einen komplementären –RNS-Strang her. Er dient als Vorlage für die Synthese vieler +RNS-Stränge in der befallenen E. coli-Zelle, die direkt als mRNS für die Synthese von Bakteriophagenproteinen dienen. In der Wirtszelle liegen jedoch viele wirtseigene RNS-Moleküle vor, welche durch die Q
-Replikase. Dieses Enzym stellt vom +RNS-Strang einen komplementären –RNS-Strang her. Er dient als Vorlage für die Synthese vieler +RNS-Stränge in der befallenen E. coli-Zelle, die direkt als mRNS für die Synthese von Bakteriophagenproteinen dienen. In der Wirtszelle liegen jedoch viele wirtseigene RNS-Moleküle vor, welche durch die Q![]() -Replikase nicht vervielfacht werden sollen – das wäre für das Virus nachteilig. Deshalb verfügt die Q
-Replikase nicht vervielfacht werden sollen – das wäre für das Virus nachteilig. Deshalb verfügt die Q![]() -Replikase über eine Erkennungsstelle, welche eine ganz bestimmte Basensequenz auf der Bakterienvirus-mRNS erkennt. Die Replikase bindet an dieser Sequenz und vervielfältigt die Virus-RNS. Die synthetisierte +mRNS wird mit Phagenproteinen umgeben und das fertige Virus verlässt die Zelle. Eine durch ein Virus infizierte Wirtszelle produziert etwa 1000 Phagenpartikel.
-Replikase über eine Erkennungsstelle, welche eine ganz bestimmte Basensequenz auf der Bakterienvirus-mRNS erkennt. Die Replikase bindet an dieser Sequenz und vervielfältigt die Virus-RNS. Die synthetisierte +mRNS wird mit Phagenproteinen umgeben und das fertige Virus verlässt die Zelle. Eine durch ein Virus infizierte Wirtszelle produziert etwa 1000 Phagenpartikel.
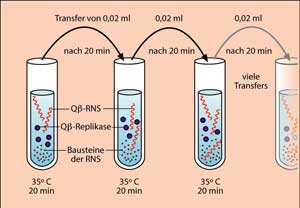
Man kann die Q![]() -Replikase des Bakterienvirus isolieren und unter den richtigen Temperatur-, Salz- und pH-Bedingungen verschiedene RNS-Moleküle im Reagenzglas vermehren. Damit eine RNS-Synthese stattfinden kann, muss man zusätzlich die vier Bausteine der RNS als Nukleotidtriphosphate in geeigneter Menge zusetzen (Abb. 1). Im folgenden werden wir ein bereits als klassisch zu bezeichnendes Experiment zur Evolution der Q
-Replikase des Bakterienvirus isolieren und unter den richtigen Temperatur-, Salz- und pH-Bedingungen verschiedene RNS-Moleküle im Reagenzglas vermehren. Damit eine RNS-Synthese stattfinden kann, muss man zusätzlich die vier Bausteine der RNS als Nukleotidtriphosphate in geeigneter Menge zusetzen (Abb. 1). Im folgenden werden wir ein bereits als klassisch zu bezeichnendes Experiment zur Evolution der Q![]() -RNS besprechen.
-RNS besprechen.
Der Aufbau des Experiments ist in Abb. 1 gezeigt. In eine Serie von Reagenzgläsern wurden unter geeigneten Bedingungen jeweils Q![]() -Replikase und Bausteine der RNS vorgelegt. In das erste Reagenzglas fügte man dann einige RNS-Moleküle mit einer Länge von 218 Nukleotiden zu und gab der Q
-Replikase und Bausteine der RNS vorgelegt. In das erste Reagenzglas fügte man dann einige RNS-Moleküle mit einer Länge von 218 Nukleotiden zu und gab der Q![]() -Replikase 20 Minuten Zeit für die Synthese von RNS-Molekülen, welche ihrerseits wieder als Matrizen (Vorlagen) zur Verfügung standen (Die Q
-Replikase 20 Minuten Zeit für die Synthese von RNS-Molekülen, welche ihrerseits wieder als Matrizen (Vorlagen) zur Verfügung standen (Die Q![]() -Replikase macht bei der RNS-Synthese immer wieder Fehler, welche zu mutierten Tochtermolekülen führen). Danach wurde eine kleine Menge des Reaktionsgemisches entnommen und in das zweite Reagenzglas überführt. Die im Schritt 1 synthetisierten RNS-Moleküle dienten jetzt als Matrizen für eine weitere Vervielfältigung der RNS. Dieser Vorgang wurde häufig wiederholt. Während eines solchen Experiments kann man verschiedene Selektionsbedingungen wählen. Beispielsweise ist bekannt, dass die Synthese von DNS und RNS durch das krebserregende Zellgift Ethidiumbromid gehemmt wird. Dieses Gift wurde zugesetzt und die Replikationsgeschwindigkeit über 20 Transfers hinweg gemessen (Abb. 2). Es zeigte sich, dass zunehmend RNS-Moleküle selektiert wurden, die immer unempfindlicher gegen Ethidiumbromid wurden. Sie konnten sich entsprechend schneller replizieren. Um die molekulare Ursache dieser Resistenz aufzuklären, wurden zu verschiedenen Zeitpunkten Sequenzanalysen der RNS durchgeführt. Dabei zeigte sich, dass drei ganz bestimmte Punktmutationen in einer bestimmten zeitlichen Reihenfolge in der Sequenz aufgetreten waren, welche die verbesserte Resistenz gegen Ethidiumbromid verursachten. Genau die gleichen Punktmutationen wurden in unabhängigen Wiederholungen des Experiments gefunden.
-Replikase macht bei der RNS-Synthese immer wieder Fehler, welche zu mutierten Tochtermolekülen führen). Danach wurde eine kleine Menge des Reaktionsgemisches entnommen und in das zweite Reagenzglas überführt. Die im Schritt 1 synthetisierten RNS-Moleküle dienten jetzt als Matrizen für eine weitere Vervielfältigung der RNS. Dieser Vorgang wurde häufig wiederholt. Während eines solchen Experiments kann man verschiedene Selektionsbedingungen wählen. Beispielsweise ist bekannt, dass die Synthese von DNS und RNS durch das krebserregende Zellgift Ethidiumbromid gehemmt wird. Dieses Gift wurde zugesetzt und die Replikationsgeschwindigkeit über 20 Transfers hinweg gemessen (Abb. 2). Es zeigte sich, dass zunehmend RNS-Moleküle selektiert wurden, die immer unempfindlicher gegen Ethidiumbromid wurden. Sie konnten sich entsprechend schneller replizieren. Um die molekulare Ursache dieser Resistenz aufzuklären, wurden zu verschiedenen Zeitpunkten Sequenzanalysen der RNS durchgeführt. Dabei zeigte sich, dass drei ganz bestimmte Punktmutationen in einer bestimmten zeitlichen Reihenfolge in der Sequenz aufgetreten waren, welche die verbesserte Resistenz gegen Ethidiumbromid verursachten. Genau die gleichen Punktmutationen wurden in unabhängigen Wiederholungen des Experiments gefunden.
|
Wir halten als Ergebnis fest, dass eine schon vorhandene Eigenschaft durch einen echten „Darwin-Prozess“ im Reagenzglas optimiert wurde. Selektionsgesteuerte Evolutionsprozesse treten unter geeigneten Bedingungen auch im zellfreien System auf. Dies spielt auch in der Diskussion der Lebensentstehung eine Rolle (Kapitel IV.7).
Das erste Beispiel für einen solchen Vorgang wurde ebenfalls schon vor längerem im Q![]() -System entdeckt. Die Q
-System entdeckt. Die Q![]() -Replikase kann erstaunlicherweise RNS auch ohne Matrize synthetisieren, weil das Protein in seinem aktiven Zentrum Bindungsstellen für einzelne Basen hat. Wenn die Q
-Replikase kann erstaunlicherweise RNS auch ohne Matrize synthetisieren, weil das Protein in seinem aktiven Zentrum Bindungsstellen für einzelne Basen hat. Wenn die Q![]() -Replikase in Gegenwart einer bestimmten (nicht zu geringen) Konzentration von einzelnen Nukleotidtriphosphaten bebrütet wird, setzt mit der Zeit eine de novo RNS-Synthese ein – es entsteht eine völlig neuartige RNS-Sequenz. Sobald derartige RNS-Matrizen vorliegen, benutzt die Q
-Replikase in Gegenwart einer bestimmten (nicht zu geringen) Konzentration von einzelnen Nukleotidtriphosphaten bebrütet wird, setzt mit der Zeit eine de novo RNS-Synthese ein – es entsteht eine völlig neuartige RNS-Sequenz. Sobald derartige RNS-Matrizen vorliegen, benutzt die Q![]() -Replikase diese als neue Vorlagen. Wenn jetzt das in Abb. 1 dargestellte Experiment durchgeführt wird, evolvieren diese anfänglichen Zufallssequenzen. Im Endstadium enthalten sie zum großen Teil die Erkennungssequenz für die Q
-Replikase diese als neue Vorlagen. Wenn jetzt das in Abb. 1 dargestellte Experiment durchgeführt wird, evolvieren diese anfänglichen Zufallssequenzen. Im Endstadium enthalten sie zum großen Teil die Erkennungssequenz für die Q![]() -Replikase, sind in ihrer räumlichen Faltung stabil und so aufgebaut, dass sie schnell durch die Replikase vervielfältigt werden können.
-Replikase, sind in ihrer räumlichen Faltung stabil und so aufgebaut, dass sie schnell durch die Replikase vervielfältigt werden können.
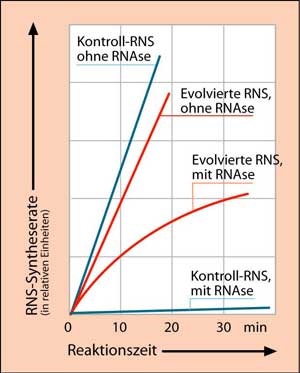
Dieses Experiment kann man auch in Gegenwart einer RNAse (ein RNS spaltendes Enzym) durchführen. Es zeigt sich, dass die unter diesem Selektionsdruck evolvierten RNS-Sequenzen eine spezielle, durch Basenpaarung bedingte Faltung (Sekundärstruktur) aufweisen, welche ihnen eine Resistenz gegen den Abbau durch die RNAse verleiht (Abb. 3).
Das Außergewöhnliche an diesem Experiment ist, dass keine RNS-Matrize als Ausgangsmaterial zur Verfügung stand. Zwar ist ein Einfluss der komplexen Q![]() -Replikasestruktur auf die entstehenden RNS-Moleküle nicht auszuschließen, doch kann man sicher sagen, dass bestimmte Strukturen mit selektionspositiven Eigenschaften neu (de novo) entstanden sind.
-Replikasestruktur auf die entstehenden RNS-Moleküle nicht auszuschließen, doch kann man sicher sagen, dass bestimmte Strukturen mit selektionspositiven Eigenschaften neu (de novo) entstanden sind.
|
Dies konnte inzwischen auch in ganz anderen Systemen gezeigt werden. Eine amerikanische Arbeitsgruppe hat 1995 erstmals die de novo Entstehung und anschließende evolutionäre Optimierung eines Ribozyms im Reagenzglas demonstriert (Abb. 4). Ein Ribozym ist ein enzymatisch aktives RNS-Molekül. Im vorliegenden Fall verfügt es über die Fähigkeit, zwei RNS-Moleküle zu verbinden (Ligaseaktivität). Im Experiment wurden Zufallssequenzen von 220 Basen Länge erzeugt. Diese Sequenzlänge lässt 4220 ![]() 10132 unterschiedliche Sequenzvarianten zu. In dem Experiment wurden nur etwa etwa 1015 Moleküle erzeugt (zum Vergleich: ein Mol eines Stoffes enthält etwa 6 x 1023 Moleküle). Dennoch waren darunter mehrere mit einer sehr niedrigen enzymatischen Aktivität. Das Experiment zeigte, dass rund ein Molekül unter 1014 Sequenzen eine Ribozym-Aktivität besaß, welche in einer anschließenden Reagenzglasevolution optimiert werden konnte.
10132 unterschiedliche Sequenzvarianten zu. In dem Experiment wurden nur etwa etwa 1015 Moleküle erzeugt (zum Vergleich: ein Mol eines Stoffes enthält etwa 6 x 1023 Moleküle). Dennoch waren darunter mehrere mit einer sehr niedrigen enzymatischen Aktivität. Das Experiment zeigte, dass rund ein Molekül unter 1014 Sequenzen eine Ribozym-Aktivität besaß, welche in einer anschließenden Reagenzglasevolution optimiert werden konnte.
Dieses Ergebnis ist bedeutsam: Es zeigt, dass eine Ribozymaktivität grundsätzlich aus Zufallssequenzen entstehen kann, weil im Sequenzraum von 10132 möglichen Sequenzen rund 10132 x 10–14 ![]() 10118 „primitive“ Lösungen vorliegen (Ob unter 10130 Proteinsequenzen auch derart viele Lösungen vorliegen, ist unbekannt, doch gibt es einige Argumente, die eher dagegen sprechen). Dieses Experiment konnte nur durch den geplanten Einsatz von komplexen Proteinen unter exakt eingestellten Laborbedingungen durchgeführt werden – von Ursuppenbedingungen ist es weit entfernt (vgl. IV.7).
10118 „primitive“ Lösungen vorliegen (Ob unter 10130 Proteinsequenzen auch derart viele Lösungen vorliegen, ist unbekannt, doch gibt es einige Argumente, die eher dagegen sprechen). Dieses Experiment konnte nur durch den geplanten Einsatz von komplexen Proteinen unter exakt eingestellten Laborbedingungen durchgeführt werden – von Ursuppenbedingungen ist es weit entfernt (vgl. IV.7).
Zusammenfassend kann man sagen, dass Evolutionsvorgänge im Reagenzglas simuliert werden können. Dabei werden vorhandene Strukturen optimiert. Bis zu einem gewissen Grad können auch neue Strukturen unter bestimmten Bedingungen erzeugt werden. Wir stehen allerdings erst am Anfang der Erforschung solcher Evolutionsvorgänge im Reagenzglas. Sie werden durch ausgefeilte molekularbiologische Techniken in immer größerem Umfang ermöglicht und werden mit Sicherheit noch wichtige Einblicke in die Struktur-Funktionsbeziehungen und damit die Wahrscheinlichkeit der Evolution von biologischen Makromolekülen liefern.