Der Artikel gibt den Stand der 6. Auflage von 2006 wieder. Eine Aktualisierung, die dem Stand im Lehrbuchtext der 7. Auflage entspricht, ist geplant. Zusammenfassung: In diesem Artikel wird die Endosymbionten-Hypothese (ESH) erklärt und es werden die Argumente erläutert, die zugunsten dieser Hypothese angeführt werden. Des weiteren werden die wichtigsten neueren Befunde zusammengestellt, die die ESH als fragwürdiger erscheinen lassen, als dies noch vor zwei Jahrzehnten der Fall war. Damit wird begründet, weshalb die ESH nicht als bewiesen gelten kann. Eine Reihe von Befunden passt besser zu einer planvollen Entstehung der Zellorganellen und ihrer Interaktionen mit dem Zellkern. Inhalt
Die gesamte Organismenwelt lässt sich in drei große Gruppen einteilen: Die Bakterien (inklusive der Cyanobakterien, früher Blaualgen), die Archaea und die Eukaryoten. Die Archaea unterscheiden sich zwar in vielen Merkmalen von den Bakterien, aber sie besitzen ebenfalls keinen echten Zellkern. Bakterien und Archaea werden daher zu den Prokaryoten zusammengefasst (Abb. 1). Eukaryoten umfassen alle übrigen Lebewesen, von Einzellern bis hin zu Pflanzen und Tieren. Der große Unterschied der Eukaryoten zu den anderen Gruppen besteht neben dem Besitz eines echten Kerns darin, dass sie Organellen enthalten, die durch Membranen vom Zellplasma abgegrenzt sind und ein eigenes Genom (= Erbgut) beherbergen: Mitochondrien und Plastiden (vgl. Abb. 2). Mitochondrien setzen Glukose und Fette in Energie um und werden daher als „Kraftwerke der Zelle“ bezeichnet. Pflanzen besitzen zusätzlich zu den Mitochondrien Plastiden. Diese lassen sich in mehrere Gruppen einteilen, darunter sind die bekanntesten die Chloroplasten. In ihnen erfolgt die Umwandlung von Sonnenenergie in biochemische Energie (Glukose) durch Photosynthese. Den Chloroplasten verdanken die Pflanzen ihre grüne Farbe (chloros = grün).
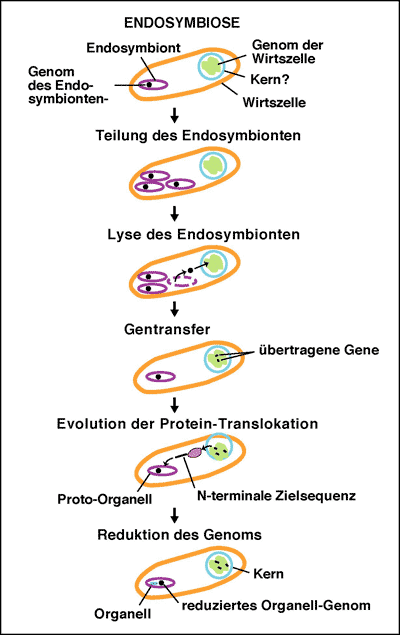
Im Rahmen der Evolutionstheorie wird vermutet, dass die Organellen in den eukaryotischen Zellen von den einfacher aufgebauten Prokaryoten abstammen. Dabei wird angenommen, dass vor mehr als 1,5 Milliarden Jahren ein vormals freilebender Prokaryot von einer anderen (organellfreien) Vorläuferzelle aufgenommen wurde, um im Laufe der Zeit zum intrazellulären Organell zu werden (Gray & Doolittle 1982). Die Aufnahme eines Prokaryoten durch eine andere Zelle soll somit die evolutive Entstehung der Eukaryoten markieren. Beide Organismen sind durch diesen Prozess eine Partnerschaft eingegangen, die anfangs allein zu beiderseitigem Nutzen war und schließlich zu gegenseitiger Abhängigkeit führte – eine Symbiose. Dieser hypothetische Entwicklungsschritt wird durch die Endosymbionten-Hypothese (ESH) beschrieben (Abb. 3). Die ESH wurde für Plastiden in ihrer ersten Form bereits 1883 von Schimper (vgl. Schimper 1885) und 1905 von Mereschkowsky aufgestellt, wurde jedoch erst seit ihrer Verbreitung durch Margulis in den 1970er Jahren bekannter (Martin et al. 2001). Diese Hypothese stellte – insbesondere in der Zeit ihrer Entstehung – eine faszinierende Idee dar und wurde mit vielen experimentellen Daten gestützt. Manche Biologen halten die Hinweise für die ESH für so überzeugend, dass sie die Endosymbiose als Beweis der stammesgeschichtlichen Entwicklung der eukaryotischen Organismen betrachten. In der folgenden Übersicht des derzeitigen Forschungsstandes wird jedoch deutlich, dass eine solche Einschätzung aufgrund der Datenlage nicht gerechtfertigt ist. Die ESH kann zwar weiterhin als eine durch viele Daten begründete Hypothese gelten (siehe dazu den Anhang „Die klassischen Argumente für die Endosymbionten-Hypothese“), es stehen ihr aber auch zahlreiche Befunde entgegen, die in der aktuellen Forschung zunehmend entdeckt werden. Dies wird in den nachfolgenden Abschnitten gezeigt.
Schon 1890 hat Altmann festgestellt, dass sich Mitochondrien (damals „Zellgranula“) ähnlich wie Bakterien anfärben lassen (Altmann 1890). Organellen sind von einer Doppelmembran umgeben, wobei die innere Membran bakterienähnliche Phospholipide (Cardiolipin) enthält, während die äußere Membran (mit Cholesterin) eher eukaryotischen Charakter hat. Die Existenz der Doppelmembran wird so interpretiert, dass die eukaryotische Membran der Wirtszelle beim Endozytose-Prozess die einwandernde prokaryotische Zelle mitsamt ihrer Membran umschlossen hat. Auch in weiteren Eigenschaften ähneln Mitochondrien den Bakterien, weshalb man ihre Verwandtschaft postulierte. So besitzen Organellen ihr eigenes Genom, das im Vergleich zu Prokaryoten zwar stark reduziert, aber wie bei diesen frei von Proteinen ist. Sie besitzen auch einen eigenen Protein-Synthese-Apparat, dessen Bestandteile (Ribosomen, tRNA, RNA-Polymerase, ribosomale RNA sowie die Codierung von Aminosäuren in einigen Codons) denen der Prokaryoten entsprechen. Die meisten Argumente für Verwandtschaft zwischen den Zellorganellen und Prokaryoten nehmen Bezug auf derartige Ähnlichkeiten (vgl. dazu die nachfolgende Auflistung). Einige Ähnlichkeiten könnten auch funktional interpretiert werden und damit auch konvergent entstanden sein. 1. Mitochondrien und Plastiden entstehen nur durch Teilung aus ihresgleichen. Die Zelle kann diese Organellen nicht neu bilden, wenn sie verlorengehen. 2. Der Besitz einer Doppelmembran könnte so interpretiert werden, dass die Membran eingewanderter Zellen beim Eindringen von der Membran der Wirtszelle umschlossen wurde. 3. Die innere Mitochondrienmembran enthält stets das Lipid Cardiolipin, das sonst nur in den Prokaryoten-Membranen auftritt. Die äußere Membran enthält dagegen wie andere Membranen der Eucyte (so werden die Zellen der Eukaryoten bezeichnet) Cholesterol, das weder in der inneren Membran noch bei Bakterien vorkommt. 4. Mitochondrien und Plastiden enthalten DNS, die wie bei Prokaryoten „nackt“, d.h. nicht mit Proteinen verbunden ist. Sie besitzen auch einen eigenen Proteinsynthese-Apparat, dessen Bestandteile (Ribosomen, tRNS und RNS-Polymerase) denen der Prokaryoten entsprechen. 5. Ribosomale RNS der Plastiden bzw. Mitochondrien hat große Ähnlichkeit mit der RNS prokaryotischer Ribosomen. 6. Mitochondrien reagieren auf manche (nicht alle) gegen Bakterien gerichtete Antibiotika. 7. Unter den heute lebenden Organismen gibt es Fälle von Symbiosen zwischen plastidenlosen einzelligen Flagellaten und Algenzellen, die als Modell für eine bestimmte Stufe des phylogenetischen Endosymbiosevorgangs angesehen werden könnten.
Über den Charakter der ursprünglichen Wirtszelle, die eine Endosymbiose einging, kann nur spekuliert werden. Bislang ist man davon ausgegangen, dass ein Prokaryot zunächst einen Zellkern entwickelt hat (eine sog. proto-eukaryotische Zelle), um in einem nachfolgenden, unabhängigen Ereignis einen anderen Prokaryoten als Proto-Mitochondrium aufzunehmen. Dieses Szenario wurde als serielle Endosymbiose bezeichnet (Gray 1999). Als Modellorganismen für einen derartigen Proto-Eukaryoten galten bis vor kurzem die Gruppe der Amitochondriatae, mitochondrienlose Protisten. Allerdings wurden inzwischen in allen Amitochondriatae Überreste von Mitochondrien gefunden, entweder als kryptische Organelle (beispielsweise als Mitosomen in einigen Amöben und menschlichen Parasiten) oder als in die Kern-DNA transferierte mitochondriale Gene (Andersson et al. 2003; Burger et al. 2003; Embley & Hirt 1998; Gray et al. 2004). Da die Amitochondriatae ihre Mitochondrien also verloren oder reduziert haben, muss von „sekundären Amitochondriatae“ gesprochen werden (Gray 1999; Lang et al. 1999a), die bislang in der ESH fälschlicherweise an den Anfang des Entwicklungs-Stammbaumes platziert wurden (Philippe et al. 2000a). Diese Erkenntnis hat die Forschung in der ESH neu belebt, weshalb viele Schlüsselpositionen im Stammbaum umgestellt werden mussten (Embley & Martin 2006). Als am wahrscheinlichsten gilt demnach mittlerweile, dass die Entstehung des Zellkerns mit entsprechendem Kerngenom zeitgleich mit der Aufnahme des Endosymbionten erfolgt ist. Neuere Untersuchungen weisen darauf hin, dass gerade der Endosymbiont substanziell zur Entstehung des Kerngenoms beigetragen hat (siehe unten). Die Anwesenheit eines Mitochondriums und die Entstehungsgeschichte der Eukaryoten sind damit untrennbar verbunden.
Die Prokaryoten lassen sich in zwei Gruppen einteilen, die Archaebakterien, die zuerst in Extrem-Biotopen (z. B. Salzseen oder heißen Schwefelquellen) entdeckt wurden, und die Eubakterien, die die Mehrzahl der Bakterien ausmachen und durch die Zellwand-Komponente Murein charakterisiert sind. Derzeit wird angenommen, dass der hypothetische Wirt der Endosymbiose den Archaebakterien ähnlich war. Ein Vorläufer des Endosymbionten hingegen wird unter den Eubakterien vermutet. Aufgrund von vergleichenden Genomdaten kommen Vertreter der alpha-Proteobakterien (sie gehören zu den Gram-negativen Eubakterien und wurden früher Purpur-Bakterien genannt) als Kandidaten für einen Endosymbionten in Frage, der sich intrazellulär zum Organell entwickelt haben könnte (Andersson et al. 1998; Gray et al. 1999; Gray 1999). Das seit 1975 als möglicher Vorläufer diskutierte alpha-Proteobakterium Paracoccus denitrificans (John & Whatley 1975) wurde dabei inzwischen von Rickettsia prowazekii abgelöst (einem intrazellulären, parasitären alpha-Proteobakterium und Erreger einer Form von Typhus). Dessen Genom weist am meisten Ähnlichkeiten zum mitochondrialen Genom von Eukaryoten auf (Andersson et al. 1998), was ihn für einen hypothetischen Endosymbionten geeignet erscheinen lässt.
Auf der anderen Seite besitzt der Eukaryot Reclinomonas americana (ein heterotropher, flagellierter Frischwasser-Protozoe) ein Mitochondrium, dessen Genom dem von Bakterien am ähnlichsten ist, was es als ein dem Symbiose-Ereignis recht nahestehendes Organell interessant macht. In der Tat besitzt R. americana das größte bisher bekannte mitochondriale Genom (67 Proteine). Gleichzeitig ist das Genom des alpha-Proteobakteriums Rickettsia prowazekii mit 834 proteincodierenden Genen sehr klein (Abb. 4). Daher stellen diese beiden Organismen den derzeit kleinsten Graben zwischen eubakteriellem und mitochondrialem Genom dar (Abb. 5; Lang et al. 1997). Das mitochondriale Genom des Eukaryoten R. americana und das prokaryotische Genom von R. prowazekii könnten also einen gemeinsamen alpha-Proteobakteriellen Vorfahren teilen. Allerdings gibt es bei dem Vergleich der beiden Genome folgende Probleme: 1. R. prowazekii hat eine für intrazelluläre Parasiten typische Genomreduktion erlebt (Andersson et al. 2003; Batut et al. 2004). 2. Die Reduktion der Genome von R. americana-Mitochondrien und R. prowazekii ist in unabhängigen Prozessen durch Anpassung an das jeweilige Habitat erfolgt. 3. Beide Genome wurden in einem frühen evolutiven Stadium und dort schnell und umfassend reduziert, was aus vergleichenden Studien des Gen-Gehalts geschlossen wird (Andersson et al. 1998; Gray et al. 1999). Die beiden Organismen dienen daher zwar als ungefähre Modelle für die ESH, bieten aber keine Grundlage für ein tatsächliches endosymbiontisches Ereignis. Die Homologien in den beiden Genomen sind nicht auffälliger als Homologien sonst innerhalb der Organismenwelt, und der Größenunterschied zwischen beiden Genomen bleibt trotz relativer Nähe signifikant (67 vs. 834 codierende Gene; Abb. 4). Außerdem vernachlässigt diese Betrachtung, dass es auch zwischen Mitochondrien verschiedener Eukaryoten erhebliche Variationen hinsichtlich Genomgröße und -organisation gibt (Burger et al. 2003; Gray et al. 2004). Genom-Vergleiche, die solche Variationen berücksichtigen, zeigen, dass das Archaebakterium Sulfolobus ssp. sowie das Eubakterium Clostridium ssp. ca. 50% mehr Ähnlichkeiten zum mitochondrialen Genom von Tieren und Pilzen aufweisen als R. prowazekii, was sie als geeignetere Kandidaten für einen Wirt bzw. Endosymbionten auszeichnet (Karlin et al. 1999). In Stammbaum-Analysen, die zu Eubakterien homologe Gene von Mitochondrien mit eubakteriellen Genen vergleichen, zweigen Mitochondrien früher von der eubakteriellen Stammlinie ab als die alpha-Proteobakterien, von denen sie aber abstammen sollen (Burger et al. 2003; Gray et al. 1999; Timmis et al. 2004). Es herrscht also weiterhin über den Wirt als auch den Endosymbionten erhebliche Unklarheit (Dyall et al. 2004; Timmis et al. 2004); eine einigermaßen widerspruchsfreie Abfolge liegt nicht vor. Der evolutionäre Graben zwischen Prokaryoten und Eukaryoten ist heute größer und die Beschaffenheit des Wirtes, der das Mitochondrium erworben hat, unklarer als je zuvor (Embley & Martin 2006).
Einige aktuelle Symbiosen zwischen freilebenden Zellen stehen Pate für die ESH. Beispielsweise tragen viele Insekten eubakterielle Symbionten, wobei die Symbiosepartner z.T. physiologisch voneinander abhängig sind (Hoffmeister & Martin 2003; Zientz et al. 2004). In Pflanzenläusen der Familie Pseudococcidae wurden sogar intrazelluläre alpha-Proteobakterien gefunden, die ihrerseits ein weiteres alpha-Proteobakterium als Endosymbionten beherbergen. All diese Fälle beziehen sich jedoch auf spezialisierte Zellen – Bakteriozyten – innerhalb eines hochspezialisierten Gewebes im Wirt (Hoffmeister & Martin 2003) und können somit nicht mit Organellen verglichen werden, die in Zellen des gesamten Organismus vorhanden sind. Auch im jüngsten Beispiel einer bekanntgewordenen Symbiose wird der Endosymbiont nicht vererbt: ein Flagellat, Hatena, nimmt anscheinend in jeder Generation erneut die Grünalge Nephroselmis auf (Okamoto & Inouye 2005).
Die Bereitstellung von Energie durch den Endosymbionten an den Wirt war das historische Hauptargument für die ESH: endosymbiontische, autotrophe Proto-Mitochondrien liefern Energie an ihren heterotrophen Wirt, Proto-Plastiden setzen Sonnenlicht in biochemische Energie um. Beiden bietet der jeweilige Wirt sicheren Lebensraum, eine eukaryotische Zelle ist entstanden. Aufgrund von neueren Ergebnissen stellt der energetische Beitrag eines hypothetischen Endosymbionten derzeit jedoch eine zentrale Schwäche der ESH dar. Ein Indiz für die Energie-Versorgung als Triebfeder für die Endosymbiose war die ATP/ADP-Translokase, die die vom Mitochondrium generierte chemische Energie in Form von ATP aus dem Mitochondrium ins Zellplasma pumpt. Doch gerade diese Funktion stellt ein Problem beim Übergang eines freilebenden Proto-Endosymbionten zum endosymbiontischen Organell dar, denn ATP/ADP-Transporter in Bakterien sind grundsätzlich nach innen gerichtet: kein freilebendes Bakterium kann ATP nach außen transportieren, um sich als attraktiven Symbiosepartner für einen heterotrophen Wirt anzubieten (Amiri et al. 2003). Dasselbe gilt für Plastiden und intrazelluläre Parasiten, die ebenfalls ATP vom Zellplasma nach innen transportieren. Das mitochondriale ATP/ADP-Transportsystem kann also nicht vom Endosymbionten abstammen (Andersson et al. 2002; Karlberg et al. 2000; Martin & Müller 1998) und somit auch keine Triebfeder für die hypothetische Endosymbiose gewesen sein. Um die ESH diesbezüglich zu stützen, muss angenommen werden, dass Mitochondrien im Anschluss an das Endosymbiose-Ereignis eine nicht verwandte, wirtcodierte, Translokase-Maschinerie erworben haben, die ATP nach außen transportieren kann, um anschließend das existierende, nach innen gerichtete System zu verlieren (Amiri et al. 2003; Andersson et al. 2002). Außerdem soll erst der eukaryotische Beitrag zur Bildung des Organellgenoms den Endosymbionten in ein ATP-exportierendes Organell verwandelt haben (Karlberg et al. 2000). Damit ist die einfachste Version der ESH, nämlich die Aufnahme eines aeroben, sauerstoffveratmenden, Energie-liefernden Symbionten ausgeschlossen (Andersson & Kurland 1998; Gray 1999; Karlberg et al. 2000; Martin & Müller 1998). Ein Ausweg aus dem Wegfall von aeroben Endosymbionten als mögliche Vorläufer schien die Entdeckung von Hydrogenosomen zu sein (Embley et al. 2003a), die als „anaerobe Mitochondrien“ anzusehen sind (Embley et al. 2003b; Martin & Müller 1998; Rotte et al. 2000). Bei der Energiegewinnung verwenden sie statt Sauerstoff (aerobe Veratmung) Protonen als Elektronenakzeptoren unter Bildung von Wasserstoff („Hydrogen“, anaerobe Fermentation). Hydrogenosomen wurden in einer Reihe von Trichomonaden, Ciliaten sowie einigen Pilzen gefunden (Embley et al. 2003a; Rotte et al. 2000). Sie verwenden andere Enzyme und ihre energetische Effizienz ist geringer, ansonsten sind sie den Mitochondrien sehr ähnlich. Allerdings besitzen Hydrogenosomen kein Genom, weshalb sie als „degenerierte“ Mitochondrien von letzteren abstammen müssen und nicht umgekehrt (Embley et al. 2003b). Die Hydrogen-Hypothese postuliert aber im Rahmen der ESH, dass ein (fakultativ) anaerobes alpha-Proteobakterium (z.B. Rhodospirillum) ohne Verwendung von Sauerstoff den ATP-Bedarf des Endosymbiose-Wirtes decken konnte (Martin & Müller 1998). Dabei bleibt jedoch unklar, warum der anaerobe Stoffwechsel-Apparat, der eine Vielzahl an strukturellen und enzymatischen Proteinen beinhaltet, zwar treibende Kraft der Endosymbiose gewesen sein soll, aber in Mitochondrien nicht mehr vorhanden ist und von einem aeroben Stoffwechsel-System abgelöst wurde. Als gemeinsame Charakterisitik der Mitochondrien bleibt letztlich weder die Atmung noch die Bereitstellung von ATP erhalten (Embley & Martin 2006). Außerdem bleibt bei allen Modellen der Energiegewinnung weiterhin offen, wie die generierte Energie an den Wirt abgegeben werden konnte. Auch Proteom(= Gesamtheit der Proteine)-Analysen geben keinerlei phylogenetische (= stammesgeschichtliche) Indizien für eine anaerobe Funktion von Mitochondrien-Vorstufen (Andersson & Kurland 1999; Kurland & Andersson 2000; Tielens et al. 2002). Alternative Erklärungen dazu vermuten einen unabhängigen Ursprung von Mitochondrien und Hydrogenosomen (Dyall et al. 2004). Die ESH kann die Symbiose eines Wirtes mit einem Energie-liefernden Endosymbionten aus energetischer Sichtweise nicht mehr erklären; gerade diese hat jedoch zur Aufstellung der Hypothese geführt (Andersson & Kurland 1998; Gray 1999; Karlberg et al. 2000; Martin & Müller 1998). Die eigentliche Grundlage der ESH ist damit verloren gegangen. Der symbiontische Aspekt muss somit nach dem Verlust des Hauptargumentes für den Organell-Erwerb mit anderen Argumenten gestützt werden.
Eine alternative Sicht zur problematischen Rolle der Bereitstellung von Energie in der ESH postuliert, dass der ursprüngliche Endosymbiont keine Energie geliefert hat, sondern als energieverbrauchender Parasit in den Wirt eingedrungen ist (Andersson et al. 2002). Eine starke Reduktion des Genoms in Organellen wie in Parasiten scheint dies zu bestätigen. Die Ursachen der Genomreduktionen unterscheiden sich zwischen Parasiten und Zellorganellen jedoch wesentlich: Während Parasiten Gene verlieren, die sie nicht mehr benötigen, da sie in einer nährstoffreichen, intrazellulären Umgebung leben, sind Organellen weiterhin auf die Funktion ihrer sämtlichen Gene angewiesen. Es ist lediglich zu beobachten, dass sie eigene Gene an den Gen-Apparat des Wirtes abgeben, um die Genprodukte von dort wieder ins Organell zurück zu importieren (Timmis et al. 2004). Rezente Parasiten – wie z.B. R. prowazekii – hingegen tragen durch ihre Genomreduktion nicht zur Zellfunktion bei, sondern ahmen nur etablierte zelluläre Interaktionen zwischen Zellplasma und Mitochondrien nach, um so vom Stoffwechsel zu profitieren. Es gibt daher keinen Hinweis auf die Rolle eines Parasiten als einen zukünftigen symbiontischen Partner (Martin & Müller 1998).
Die Herkunft der Plastiden scheint klarer zu sein als der Ursprung der Mitochondrien. Hier werden cyanobakterienähnliche Vorläufer als Endosymbionten angenommen, die als „kleine grüne Sklaven“ in einer Wirtszelle Sonnenlicht in biochemische Energie umwandelten (Abb. 3; Dyall et al. 2004). Bei der vermuteten Endosymbiose von Plastiden wird zwischen zwei Symbiose-Ereignissen unterschieden: In der primären Endosymbiose nimmt eine eukaryotische Zelle, die bereits mit einem Mitochondrium ausgestattet ist, einen Prokaryoten (z.B. ein Cyanobakterium) auf, der photosynthetisch aktiv ist und zum Plastid wird (Abb. 6). Diese primären Plastiden sind – wie Mitochondrien – mit einer Doppelmembran ausgestattet. Der eukaryotische Wirt wurde dadurch autotroph, was zur Entstehung der grünen Landpflanzen, der Rotalgen und der Glaukophyten (einer kleinen Gruppe von Algen) geführt haben soll. Alle anderen photosynthetischen Eukaryoten (Euglenophyten und Chlorarachniophyten einerseits, Dinoflagellaten, Heterokonta, Haptophyten, Apicomplexa und Kryptomonaden andererseits) sollen ihre Plastiden durch eine sekundäre Endosymbiose erhalten haben. Eine eukaryotische Alge (ihrerseits schon mit Mitochondrium und Plastid) wird von einem eukaryotischen Wirt aufgenommen, so dass vorübergehend ein Eukaryot im Eukaryot lebt (Abb. 6; Kroth 2002; Moreira & Philippe 2001). Im Anschluss soll der Endosymbiont Zellplasma-Bestandteile verloren und den Zellkern zum Nucleomorph reduziert haben, um zu einem „second-hand"-Chloroplasten zu werden. Als Folge sind sekundäre, komplexe Plastiden mit mehr als zwei Plastiden-Membranen umgeben (3-4 oder auch mehr; Kroth 2002). Die photosynthetische Sauerstoffproduktion der sekundären Plastiden übertrifft die aller Plastiden von Landpflanzen, weshalb sie von großer ökologischer Bedeutung sind. Im Gegensatz zur primären Endosymbiose, die in einem einzigen Ereignis stattgefunden haben soll, werden mehrere unabhängige sekundäre Endosymbiosen angenommen (Kroth 2002). Die Sequenzierung des Genoms der thermophilen Rotalge Cyanidioschizon merolae zeigt inzwischen jedoch, dass ein Großteil der Protein-Import-Komplexe zwischen beiden Plastidenformen konserviert ist, was einen ersten Hinweis auf einen monophyletischen Ursprung aller Plastiden gibt (McFadden & van Dooren 2004). Sowohl bei primären als auch sekundären Endosymbiosen sind Wirt und Plastid molekular und physiologisch komplex miteinander verknüpft und nur so funktionell. Eine evolutive Entwicklung dieses Netzwerkes durch natürliche Selektion muss allerdings als sehr unwahrscheinlich betrachet werden (siehe Abschnitt „Sortierung und Import von Proteinen“).
Die Betrachtung des Genoms der Mitochondrien unter dem Versuch, das historische Ereignis der Endosymbiose nachzuvollziehen, ergibt ein komplexes Bild: Im Anschluss an die Aufnahme eines Endosymbionten muss eine drastische Reduktion des Organell-Genoms stattgefunden haben. Dabei sind ein paar hundert (ein Großteil) der Gene eines ursprünglichen Endosymbionten verloren gegangen (v.a. solche, die energetische und Translations-Prozesse unterstützen). Das Proto-Mitochondrium wurde schließlich so in den Zellstoffwechsel eingebunden, dass es an nahezu allen physiologischen Vorgängen beteiligt ist. Der hypothetische Endosymbiont hat also auf diese Weise seine Autonomie sowie seine eubakterielle Identität völlig verloren, denn nur zwischen 14-20% der mitochondrialen Proteine sind auf eine alpha-proteobakterielle Herkunft zurückzuführen (Abb. 7; Karlberg et al. 2000). Die Zusammenstellung der mitochondrialen Proteine aus einer kleinen Gen-Fraktion des alpha-proteobakteriellen Endosymbionten und einer größeren Fraktion eukaryotischer Gene wird in der „dual origin hypothesis“ beschrieben (Karlberg et al. 2000). Das Genom der Eukaryoten selbst scheint von der Genomreduktion des hypothetischen Endosymbionten durch Übernahme von Genen profitiert zu haben, denn das Genom des Symbionten soll einen „riesigen Pool“ an Genen dargestellt haben (Abb. 7). Der Erwerb eines mitochondrialen Endosymbionten soll eine entscheidende Bedingung für die Entstehung der Eukaryoten gewesen sein; jedenfalls wird beides inzwischen als Teil desselben Ereignisses betrachtet (Lang et al. 1999b). Der proto-mitochondriale Gentransfer hat gewissermaßen die Eukaryoten aus der Gruppe der Prokaryoten herausgeschmiedet (Timmis et al. 2004), und die Aufnahme eines Endosymbionten wird daher als Hauptursache der Eukaryogenese (Entstehung der Eukaryoten) gedeutet (Herrmann et al. 2003; Lang et al. 1999a). In der weiteren Evolution muss der Gentransfer zwischen den verschiedenen Wirten und ihren Organellen sehr unterschiedlich verlaufen sein, denn von den 150 bis 4000 mitochondrialen Proteinen sind nur etwa 50 in sämtlichen Eukaryoten universal vorhanden (Timmis et al. 2004). Die Integration der Organelle in nahezu alle regulatorischen und metabolischen Zellvorgänge verlief also – artspezifisch – so spezialisiert, dass der Vergleich heutiger Organellen eine hochkomplexe Restrukturierung und Mischung der ursprünglichen, endosymbiontischen Partner aufzeigt. Die Unterschiede sind z.T. sogar so groß, dass einige Biologen annehmen, dass Mitochondrien in der Evolution mehrmals (konvergent) entstanden sein müssen (Burger et al. 2003).
Auch wenn die Organell-eigene DNA nur für wenige Gene codiert, bestehen Mitochondrien und Plastiden aus vielen hundert Proteinen. Bei der mitochondrialen Genom-Reduktion sind demnach nicht nur Gene verloren gegangen, sondern ein Teil des genetischen Materials wurde vom hypothetischen Endosymbionten in das Kerngenom der eukaryotischen Wirtszelle transferiert (Abb. 7; Dyall et al. 2004). Dieses hypothetische Szenario des massiven Gentransfers ist in Abb. 8 dargestellt. Es wird vermutet, dass das Organell-Genmaterial entweder durch die gelegentliche Lyse ganzer Mitochondrien oder Chloroplasten („bulk DNA“) oder durch DNA-Zwischenprodukte ins Kerngenom gelangt ist (Stegemann et al. 2003). Der Gentransfer in den Kern hat zur Folge, dass Organell-Proteine nun von der Kern-DNA transkribiert und im Zellplasma translatiert werden müssen, um in die Organellen zurücktransportiert zu werden. Es wird vermutet, dass eine Lokalisierung der Organell-Gene im Kerngenom Vorteile hat, wie z. B. eine geringere Frequenz schädlicher Mutationen durch sexuelle Rekombination und eine räumliche Trennung der DNA von den Sauerstoff-Radikale produzierenden biochemischen Prozessen in den Organellen (Martin & Herrmann 1998). Lediglich Proteine, die entweder zu hydrophob sind, um effizient in die Organellen transportiert zu werden, oder solche, die rasch auf Änderungen im Redox-Potenzial der Organellen reagieren müssen, wurden im Organell-Genom behalten (Race et al. 1999). Aus Experimenten, in denen Transfer von Genen aus mitochondrialer (in der Bäckerhefe Saccharomyces cerevisiae) bzw. plastidärer DNA (in der Tabakpflanze Tobacco ssp.) ins Kerngenom gezeigt wurde, wird gefolgert, dass solche Gentransfer-Ereignisse auch aktuell stattfinden (Shafer et al. 1999; Thorsness et al. 1993). Inzwischen wurden Vorgänge beobachtet, durch welche manche mitochondrialen Gene offensichtlich mehrmals unabhängig voneinander ins Kerngenom kopiert wurden bzw. Gene in zwei Hälften an verschiedene Loci im Genom transferiert wurden, wobei die Proteinprodukte nun posttranslational fusionieren müssen (Adams & Palmer 2003; Burger et al. 2003). Manche Plastiden haben im Verlauf dieser Genom-Reduktion sogar ihre photosynthetische Funktion verloren, wie die Apicoplasten – von Plastiden abgeleitete Organellen einer Gruppe von menschlichen Parasiten, zu denen auch der Malaria-Erreger Plasmodium ssp. gehört (Martin & Borst 2003; Waller et al. 2004). Ungenügend untersucht ist jedoch, ob in einem Wirt von Endosymbiont zu Kerngenom transferierte Gene auch zwischen verschiedenen Spezies gleich organisiert sind und demnach auseinander hervorgegangen sein können, wie dies für den Gentransfer im Rahmen der ESH gefordert wird.
Neben der Reduktion des Organell-Genoms und dem Transfer mitochondrialer Gene in das Kerngenom des Wirtes mussten dort hingegen ein paar weitere hundert Gene neu entwickelt oder erworben worden sein. Insbesondere Gene, die regulatorische und Transportfunktionen garantieren, mussten neu in den Dienst des sich entwickelnden Organells gestellt werden, damit die transferierten Gene überhaupt genutzt werden konnten (Abb. 7; Andersson et al. 2003; Burger et al. 2003; Dyall et al. 2004; Karlberg et al. 2000). Diese Gene unbekannter Herkunft codieren immerhin einen Großteil des mitochondrialen Proteoms. Sie sollen entweder völlig neu durch Spontanmutationen entstanden oder von anderen ebenfalls unbekannten Organismen durch lateralen Gentransfer (Austausch von Genen zwischen verschiedenen Organismen) erworben worden sein. Die meisten kerncodierten, mitochondrialen Proteine zeigen jedenfalls keine Homologien zu heutigen bakteriellen Proteinen (Andersson et al. 2002). Daher kann nur ein Teil dieser unbekannten Gene auf lateralen Gentransfer zurückgeführt werden. Letzterer wird ohnehin in der Erklärung von Homologien überstrapaziert (Martin 2003; Timmis et al. 2004). Für den Großteil der mitochondrialen Proteine muss demnach eine de novo Enstehung angenommen werden – ein sehr unwahrscheinliches Szenario, das außerdem in evolutiv kürzester Zeit geschehen sein muss (vgl. Abschnitt „Phylogenetische Analysen“).
Eine grundlegende Konsequenz des Massentransfers von Organell-Genen in das eukaryotische Kerngenom ist die Notwendigkeit, die im Kern codierten mitochondrialen und plastidären Proteine posttranslational zurück in das Organell zu transportieren. Dieser komplexe Vorgang erfolgt in mehreren Schritten: zunächst müssen sogenannte Zielsequenzen (20-60 Aminosäuren mit einer Zielinformation vor der Sequenz des eigentlichen Proteins, Abb. 8) ein Protein zum richtigen Organell und dort an die richtige Stelle führen. Dafür werden Chaperone (Hilfsproteine) benötigt, die Proteine im Zellplasma entfaltet halten können. Anschließend müssen die kerncodierten Proteine in das Organell eingefädelt werden, was durch Translokasen (Abb. 9; hochregulatorische und -spezifische Porenkomplexe aus vielen Protein-Untereinheiten) vermittelt wird (Pfanner & Geissler 2001). An ihrem Zielort spalten dann spezifische Proteasen (Proteine, die andere Proteine schneiden können) die Zielsequenzen ab. Die importierten Proteine werden gefaltet, mit anderen Untereinheiten zu Komplexen gruppiert und ggf. in eine Membran eingebaut (Neupert 1997; Pfanner & Geissler 2001). Schließlich müssen Proteasen während des gesamten fehleranfälligen Vorgangs eine Qualitätskontrolle durchführen (Luzikov 2002). Diese Prozesse verlaufen unter Verbrauch von biochemischer Energie. Der ursprüngliche Transfer der Organell-Gene in das Kerngenom konnte nur dann erfolgreich sein, wenn diese jeweils hinter einer Zielsequenz und einer entsprechenden Promotor-Region im Genom zu landen kamen. Nur wenn ins Kerngenom kopierte Gene exprimiert und zum Organell transportiert, importiert, von ihrer Zielsequenz befreit und dort zusammengefügt wurden, war ein funktioneller Gentransfer gewährleistet (Martin 2003; Timmis et al. 2004). Da DNA-Moleküle bei der Genomreduktion zufällig in das Kerngenom eingebaut wurden, muss der Transfer eines jeden Gens als sehr unwahrscheinlich betrachtet werden. Die ESH behilft sich mit dem Argument, dass Gene als Duplikate aus dem Endosymbionten ausgewandert sind, um dort erst verloren zu gehen, nachdem ihre Kern-inserierten Kopien funktionell geworden waren. Im Falle der sekundären ESH mussten die meisten Gene sogar eine weitere Präsequenz erhalten, damit „zweiteilige Zielsequenzen“ entstehen konnten, und ein zweites Mal funktionell transferiert werden. Wie Plastiden-Gene hinter entsprechende Zielsequenzen gesetzt werden konnten, ist weitgehend unklar (Kilian & Kroth 2004;Kroth 2002). Translokasen sind große Proteinkomplexe, die aus vielen membranständigen und löslichen Untereinheiten bestehen. Mitochondrien besitzen in ihrer äußeren Membran eine Translokase, den TOM-Komplex, in ihrer Innenmembran zwei, die TIM-Komplexe (translocase of the outer/inner membrane, Abb. 9; Neupert 1997; Pfanner & Geissler 2001). In Chloroplasten heißen die entsprechenden Translokasen TOC- und TIC-Komplexe (translocase of the outer/inner chloroplast envelope, Abb. 9). Für kaum eine Untereinheit der eukaryotischen Translokasen wurden Homologe in Prokaryoten gefunden (Kroth 2002). Auch die Translokasen von Mitochondrien und Plastiden untereinander weisen keine Sequenz-Verwandtschaft auf, sondern haben nur homologe Funktionen. Da es nur Systeme mit funktioneller Ähnlichkeit gibt, wird in der ESH angenommen, dass die Translokasen entweder aus entfernt verwandten Membran-Transport-Kanälen oder de novo entstanden sind (Dyall et al. 2004; Schnell 2000; Schnell & Hebert 2003). Zusätzlich zu den Translokasen gibt es Sortierungsapparate, die für eine korrekte Insertion von Proteinen in der Membran zuständig sind (Abb. 9). Für den Oxa1 genannten Komplex in Mitochondrien (Herrmann 2003) wurde ein eubakterieller Vorläufer gefunden. In Plastiden gibt es weitere Membrantransport-Systeme, die zwischen den unterschiedlichen Subkompartimenten (Stroma und Lumen) vermitteln. Für den „cpSec-Weg“ (Protein-Import aus dem Stroma in das Lumen) und den „cpDpH-Weg“ (evtl. von einem prokaryotischen Ionen-Kanal adaptiert) kann eine homologe Funktion der Sekretion in Bakterien hergestellt werden, zwei weitere Transportwege (der „cpSRP“ und der „spontane Insertions-Weg“) haben offensichtlich keine prokaryotischen Vorläufer (Schnell 2000). Die enorme Komplexität und Spezifität dieser Protein-Komplexe macht eine solche Neuentwicklung jedoch höchst unwahrscheinlich, nicht zuletzt, weil die gruppierten Untereinheiten nur in ihrer physiologischen Vernetzung funktionell und nur dann selektiv vorteilhaft sind. Ein funktioneller Gentransfer für jedes einzelne Gen durch zufällige Transferereignisse und Selektion muss sehr lange evolutionäre Zeiträume annehmen. Auch die Annahme eines gerichteten Gentransfers hinter vorbereitete Promotor- und Zielsequenzen muss mit geringen Wahrscheinlichkeiten rechnen, bis ein Protein funktionell ins Organell zurückimportiert wird. Dabei muss nicht nur der Gentransfer jedes Gens, sondern der gesamte Vorgang der Proteinsortierung und des Proteinimports von Anfang an funktionsfähig sein, damit Selektion als positiver Evolutionsmechanismus wirken kann. Die beschriebenen Vorgänge werden alle durch die Aktion von hochkomplexen Proteinmaschinerien und unter Energieverbrauch vermittelt. Die Neuentstehung von Genen und der zufällige Einbau von Organell-Genen ins Kerngenom muss zudem aufgrund der phylogenetischen Daten in kürzester Zeit stattgefunden haben. Als alternatives Entstehungsmodell im Rahmen der Schöpfungslehre kann man daher auch eine vorbereitete Situation annehmen, in der Gene intentional eingebaut wurden. Somit könnte für jeden Organismus die spezifische, zelluläre Interaktion zwischen Wirt und Organell von Anfang funktionell sein. Eine solche Situation ist evolutiv dagegen kaum verständlich zu machen. Es werden offenkundig zu viele komplexe Proteine und ihre Steuerung en bloc benötigt, um deren Entstehung durch bekannte Evolutionsmechanismen plausibel machen zu können. Denn Selektion kann erst greifen, wenn eine Funktion gegeben ist.
Aus vergleichenden Genomanalysen kann evolutionstheoretisch geschlossen werden, dass der unidirektionale Gentransfer vom Endosymbionten in das Kerngenom in „häufigen und massiven“ Ereignissen, „schnell und umfassend“ sowie „gleichzeitig“ und „relativ kurzfristig“ stattgefunden haben muss. Entsprechend wird die Evolution der Eukaryoten als Ereignis beschrieben, in dem eine Diversifizierung schnell und zeitgleich geschehen ist (Cavalier-Smith 2000; Gray et al. 1999; Gray 1999; Lang et al. 1999b; Philippe et al. 2000a; Philippe et al. 2000b). Das heißt, dass über die Verwandtschaftsverhältnisse am Ursprung des phylogenetischen Baumes keine Aussage möglich ist und die bekannten Eukaryoten mit ihren Mitochondrien praktisch zeitgleich erscheinen, womit mehrere Symbiosen unabhängig voneinander nahezu zeitgleich abgelaufen wären. Die Eukaryogenese soll in einer „explosive quantum evolution“ bzw. in einem „big bang“ vonstatten gegangen sein, weshalb alle Stammbaum-Rekonstruktionen inzwischen stammlos skizziert werden müssen (Abb. 10). Derzeit kann kein „primitiver Eukaryot“ ausgemacht werden, womit die Frage nach dem Ursprung der Eukaryoten offen bleibt (Philippe et al. 2000a). Embley & Martin (2006) sprechen von einem „Verlust historischer Signale“, der eine Auflösung des Stammbaumes an der Wurzel unmöglich macht. Da also alle Eukaryoten mehr oder weniger gleich alt sind, müssen die Phylogenien eine Auslöschung aller hypothetischen, primitiven Formen postulieren (Abb. 10). Weder eine massive, umfassende und zeitgleiche Genomreduktion noch eine explosive Quantensprung-Evolution lassen sich aber mit einer langsamen, schrittweisen Entwicklung eines Endosymbionten vereinbaren. Diese Erklärungsnöte ergeben sich zwangsläufig, wenn auf einem gemeinsamen Ursprung aller Mitochondrien in einem einzigen Endosymbiose-Schritt beharrt wird. Eine alternative Deutung im Rahmen der Schöpfungslehre würde einen artspezifischen Einbau von Mitochondrien bzw. Plastiden in die eukaryotischen Grundtypen annehmen und die gleichzeitige Entstehung sämtlicher eukaryotischer Linien ohne Vorläufer erklären. Darüber hinaus sind nach schöpfungstheoretischer Deutung der Datenlage die hochdiversifizierten Mitochondrien und Plastiden als eine artspezifischen Zusammenstellung des Genoms und Proteoms verstehbar. Organellen können als Module unter Verwendung von Funktionseinheiten in den unterschiedlichen Grundtypen eingebaut worden sein.
Die Endosymbiontenhypothese (ESH) erlebte im Laufe der vergangenen Jahrzehnte einen erheblichen Wandel in ihren stützenden Argumenten. Mechanismen, die lange Zeit plausibel erschienen, verloren durch fortlaufende Forschung ihre Aktualität. Darunter fallen zentrale Argumente wie die serielle Endosymbiose (da es doch keine Amitochondriatae gibt), die Entwicklung eines Zellkernes vor der Aufnahme eines Endosymbionten, sowie die Bereitstellung von Energie als Initiator der Endosymbiose. Der Wegfall des letzteren als Triebfeder stellt eine deutliche Schwächung der ESH dar. Die Hauptargumente für Verwandtschaft von Prokaryoten und Organellen basieren weiterhin auf Ähnlichkeit. Obwohl in den letzten zwei Jahrzehnten zu den morphologischen und biochemischen Markern eine große Zahl vergleichender Genom- und Proteom-Daten hinzugekommen sind, wurden auf der Suche nach Vorläufer-Kandidaten für einen hypothetischen Wirt (vielleicht ein Archaebakterium) und einen Endosymbiont (vielleicht ein alpha-Proteobakterium) kaum Fortschritte erzielt – es muss weiterhin nach dem „mysterious symbiont“ gesucht werden (Timmis et al. 2004). Der derzeit kleinste Graben zwischen mitochondrialem Genom einerseits (R. americana) und prokaryotischem Genom andererseits (R. prowazekii) ist immer noch erheblich. Dieses Tandem hat zudem den Nachteil, dass es mit R. prowazekii einen Parasiten einbezieht, dessen intrazelluläre Rolle mit der eines Endosymbionten nicht vergleichbar ist. Embley & Martin (2006) formulieren, dass der evolutionäre Graben zwischen Prokaryoten und Eukaryoten derzeit tiefer und die Beschaffenheit des Endosymbionten, der ein Mitochondrium aufgenommen hat, unklarer sind als je zuvor. Im Gegenzug etabliert sich zunehmend die Sicht, dass die Entstehung der Eukaryoten und das Auftreten von Mitochondrien als Organellen Teil desselben Ereignisses sind. Die Existenz von Mitochondrien führte also zur Eukaryogenese, während der etliche Gene von einem Proto-Mitochondrium in ein Wirtgenom transferiert worden sein sollen, deren Genprodukte anschließend erfolgreich in das entstandene Organell zurücktransportiert werden müssen. Darüberhinaus muss jedoch eine Vielzahl neuer Gene entstanden sein. Für beide Prozesse sind die Wahrscheinlichkeiten äußerst gering, wenn der Mechanismus mit de novo-Synthese durch Spontanmutationen sowie mit lateralem Gentransfer und anschließender natürlicher Selektion erklärt werden soll. Desweiteren muss anhand der Unterschiede zwischen Mitochondrien angenommen werden, dass nach der Endosymbiose der Gentransfer von einem ursprünglichen Endosymbionten in das Kerngenom des Wirtes in den verschiedenen Spezies sehr unterschiedlich verlaufen ist, was durch ein einmaliges Endosymbiose-Ereignis kaum zu erklären ist. Entgegen sämtlicher Erwartungen der ESH ergibt die aktuelle Menge an unterschiedlichen Daten keine genaueren Stammbäume als früher, und die Entstehung von Mitochondrien ist in ihren Details heute weitaus weniger verstanden als sie in den 1990er Jahren zu sein schien (Gray et al. 2004; Philippe et al. 2000a). Es gibt derzeit mehr gute Theorien für die Entstehung der Eukaryoten als gute Daten (Embley & Martin 2006). Aus sämtlichen phylogenetischen Sequenzvergleichen folgt, dass die endosymbiosebedingte eukaryotische Entwicklung in gleichzeitigen, umfassenden und schnellen Prozessen vonstatten gegangen sein muss. Allerdings geben die unterschiedlichen Analyse-Methoden keine übereinstimmenden Ergebnisse, weshalb die Verwandtschaft der eukaryotischen Hauptgruppen unklar bleibt (Embley & Martin 2006). Gleichzeitig stellt sich jedoch auch immer mehr eine enorme Komplexität der physiologischen Zusammenhänge zwischen Eukaryoten-Zelle und Organellen heraus. Letztere sind artspezifisch unterschiedlich und dort in nahezu alle Lebensprozesse eingebunden. Für die Entstehung der Komplexität müssen daher ebenfalls zunehmend geringere Wahrscheinlichkeiten angenommen werden, was in der wissenschaftlichen Diskussion immer vorausgesetzt, aber selten explizit erwähnt wird. Es überrascht, dass trotz der aktuellen Daten- und Ergebnis-Situation, trotz der Widersprüchlichkeit vieler Resultate und trotz der Verlustes wesentlicher Schlüssel-Argumente viele Autoren in Nebensätzen die ESH als grundsätzlich bewiesen annehmen. Es scheint, als ob die hier zusammengestellten Ergebnisse der ESH-Literatur und deren Schlussfolgerungen nicht wahrgenommen werden. Vielmehr fungiert die Evolutionsanschauung offenbar als Voraussetzung für alle Deutungen, und ein kritisches Hinterfragen dieser Voraussetzung scheint nicht zulässig zu sein. Einer der Spezialisten für mitochondriale Evolution schreibt: „...the acquisition of mitochondria must in retrospect be viewed as a wonderful success. However, an examination of modern mitochondrial functions may not necessarily tell us what kind of relationship initiated the symbiotic event. Certainly, it was not based on the prospect of creating large multicellular organisms in a distant future“ (Andersson et al. 2003). Warum dies so sicher sein soll, wird in den Publikationen jedoch nicht erläutert, und vielleicht ist die Beschreibung „wonderful" zutreffender als vom Autor beabsichtigt. Als bewiesen gelten – wie dies zumeist in Einführungen von Publikationen anklingt – kann die ESH derzeit jedenfalls nicht. Eine zur ESH alternative Interpretation der Datenlage im Rahmen der Schöpfungslehre geht davon aus, dass Eukaryoten – inklusive ihrer Organellen – als voll funktionsfähige Einheiten von Anfang an existieren. Zellbestandteile, Zellorganellen und physiologische Funktionen wurden modulartig in unterschiedlichen Organismen kombiniert, was Ähnlichkeiten zwischen pro- und eukaryotischen Zellen bedingen könnte. Dabei sind die Organellen entsprechend der verschiedenen Grundtypen in ein komplexes Netzwerk von Zellvorgängen eingebunden. Auch die genetische Lokalisierung der Organell-Proteine im eukaryotischen Genom ist artspezifisch in der jeweiligen Zellphysiologie funktionell vernetzt. Während die von der ESH z.T. mehrmalig geforderte Makroevolution von Wirt und Endosymbionten hier als zu unwahrscheinlich abgelehnt wird, ist unumstritten, dass mikroevolutive Prozesse innerhalb der Grundtypen für weitere Diversifikation gesorgt haben. Dies kann zu Reduktionen und Degeneration, aber auch zu Weiterentwicklung und Anpassung an Änderungen der Lebensbedingungen führen, wie z.B. experimentell zu beobachtetende Gentransfers vom Organell ins Kerngenom innerhalb derselben Spezies zeigt. So würde eine Erschaffung von eukaryotischen Grundtypen die schnelle und gleichzeitige Entstehung der Eukaryoten in den phylogenetischen Analysen erklären, auf die die stammlosen Phylogenien hinweisen, inklusive der angeblichen Auslöschung aller hypothetischen, primitiven Eukaryoten-Formen. Auch das völlige Fehlen sämtlicher hypothetischer primitiver Vorstufen von Wirt und Endosymbiont wäre damit verständlich. Die Erschaffung von eukaryotischen Grundtypen kann gleichbedeutend sein mit den Szenarien, die mit „explosive quantum evolution“ oder „big bang“ umschrieben werden. Auch wenn die ESH weiterhin in einigen Aspekten einen guten und interessanten Erklärungsansatz bietet, weist sie doch erhebliche Schwächen auf und ist nicht als die einzig denkbare Hypothese zu verstehen. Gregor Wagner
| |||||||||||||||||||||||||||||||||||||||||||||||||
|
Studiengemeinschaft WORT und WISSEN e.V.
|