An dieser Stelle finden Sie über den Inhalt des Buchs „Evolution - ein kritisches Lehrbuch“ hinaus einen ausführlicheren Text zum Kapitel 14.6 „Der Ursprung der Säugetiere“.
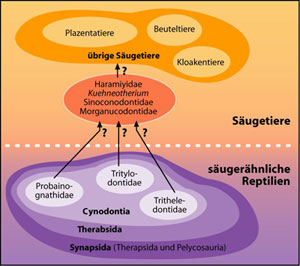
Die Säugetiere werden von den Reptilien abgeleitet. Dieser Übergang wird heute von den Evolutionsbiologen übereinstimmend als die fossil am besten dokumentierte Entstehung einer neuen großen taxonomischen Einheit angesehen. Eine genaue Betrachtung der einzelnen Formen dieses Übergangs zeigt aber große morphologische Lücken zwischen den vermuteten Bindegliedern. Zunächst muss festgestellt werden, dass wesentliche Merkmale der Säugetiere an Fossilien nicht oder nur mit einem hohen Grad an Spekulation erkannt werden können. Dazu gehören die Funktion des Herzkreislaufsystems und der Nieren, Haare, Milchdrüsen und die Temperaturphysiologie. Es ist daher ein schwieriges Unterfangen, die Evolution der Säugetiere anhand von Fossilfunden nachzuzeichnen. An Fossilien lassen sich der Bau des Schädels und des Kiefers, die Struktur der Zähne und die Skelettmerkmale des Postcranium (Körperstamm und Gliedmaßen) nachweisen. Daraus kann die Fortbewegungsweise sowie der Kau- und Hörapparat bedingt rekonstruiert werden. Unter den Reptilien gelten die sog. Synapsiden als Ahnen der Säugetiere (Abb. 5). Diese Reptilgruppe, auch „säugerähnliche Reptilien“ genannt, hat sich nach evolutionstheoretischen Vorstellungen schon fast an der Wurzel von den übrigen Reptilien abgezweigt. Sie ist heute ausgestorben. Die Beziehungen der Säugetiere zu den heute lebenden Reptilien werden daher als außerordentlich fern angesehen. Die Synapsiden sind durch ein einzelnes Fenster (eine Öffnung) im unteren Schläfenbereich an jeder Schädelseite charakterisiert (Abb. 1). Sie werden traditionell in Pelycosaurier (primitive Formen) und Therapsiden (entwickelte Formen) unterteilt.
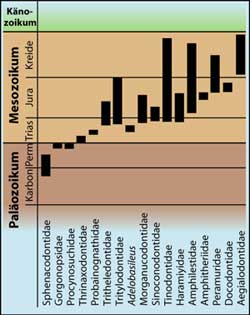
Von den Pelycosauriern über die primitiven und fortschrittlichen Therapsiden zu den frühen Säugetieren hin kann eine Reihe mit Formen aufgestellt werden, die mit zunehmend jüngerem Alter eine größere Säugetierähnlichkeit aufweisen (Abb. 2). Diese Tatsache ist das paläontologische Haupt-argument für die Vorstellung einer evolutionären Entstehung der Säugetiere aus reptilien-ähnlichen Vorfahren. Die unmittelbaren Ahnen der Säugetiere werden unter den fortschrittlichsten säuger-ähnlichen Reptilien, den Cynodontiern (Abb. 5), gesucht. Die Tritylodontidae, Tritheledontidae (Ictidosaurier) und Probainognathidae sind drei Cyno-dontierfamilien, die als besonders nahe Verwandte der Säugetiere eingestuft werden (Abb. 5). Sie sind nicht durch Zwischenformen überbrückt. Das Auftreten dieser und anderer im Text genannter Formen in der Erdgeschichte ist in Abb. 3 (auf Familienebene) dargestellt.
Als wichtigste frühe Säugetiergruppe wird die Familie der Morganucodontidae angesehen. Ausschlaggebend für die Identifikation dieser Familie als Säugetiergruppe ist der Besitz eines Kiefergelenks zwischen dem Squamosum (Schädelknochen) und dem Dentale (Unterkieferknochen), das bei allen Säugetieren, nicht aber bei den Reptilien vorkommt (Abb. 4). Unter den Wissenschaftlern bestehen jedoch bis heute unterschiedliche Auffassungen dar-über, welche fossilen Merkmale ein Säugetier charakterisieren. Da alle Reptilien außer dem Dentale weitere Unterkieferknochen besitzen, wäre der Nachweis nur eines einzigen Unterkieferknochens (Dentale) als entscheidendes Säugetierkriterium auch denkbar. Von den Morganucodontiden (Abb. 6) sind tausende Zähne, Kiefer-, Schädel- und -postcraniale Knochen vorhanden. Damit stellt diese Gruppe einen Sonderfall unter den frühen Säugetiergruppen dar. Von anderen nach evolutionären Vorstellungen wichtigen frühen Säuger-Übergangsstufen wie Kuehneotherium (Familie Tinodontidae), Amphitherium (Familie Amphitheriidae), Peramus (Familie Peramuridae) und Aegialodon (Familie Aegialodontidae) sind nur Kieferfragmente und Zähne bekannt. Da auch von den fortschrittlichen säugerähnlichen Reptilien nur zum Teil Überreste vom Körperstamm- und Gliedmaßenskelett vorhanden sind, ist der Kauapparat die fossil am besten dokumentierte Struktur im vermuteten Reptil-Säuger-Übergang. Die Morganucodontiden besitzen viele Merkmale, die auch bei den fortschrittlichen Cynodontiern zu finden sind. Darüber hinaus existiert aber auch eine Anzahl wichtiger Unterschiede. In Tab. 13.3 sind Merkmale aufgeführt, die bei allen Säugetieren, aber keinem Cynodontier zu finden sind. Wenn man nun noch gesondert die einzelnen späten Therapsidenfamilien mit den frühen Säugetieren vergleicht, dann sind die morphologischen Unterschiede weit größer. Im folgenden werden einige wichtige Gemeinsamkeiten und Unterschiede zwischen den bereits genannten Therapsidenfamilien Tritheledontidae, Tritylodontidae und Probainognathidae und der frühen Säugerfamilie Morganucodontidae aufgeführt.
Ein Hauptgrund für die Deutung von Probainognathus (einzige Gattung der Cynodontierfamilie Probai-nognathidae; Abb. 5) als eine dem Säugetierursprung nahestehende Form ist die Tatsache, dass Probainognathus von allen Cynodontiern die morganucodon-tiden-ähnlichsten Backenzähne besitzt. Andererseits gibt es aber zusätzlich zu den schon in Tab. 1 aufgeführten, alle Cynodontier betreffenden Unterschieden weitere morphologische Differenzen zu den Morganucodontiden. Dazu gehören u. a. eine fehlende Differenzierung der Backenzähne in Prämolaren und Molaren und der Besitz eines prä-frontalen und eines postorbitalen Knochens (Abb. 7). Letzterer grenzt gegenüber der Augenhöhle ein Schläfenfenster ab. Bei den Morganucodon-tiden gibt es infolge einer fehlenden postorbitalen Knochenstrebe nur eine Öffnung in diesem Bereich (siehe Abb. 6B). Ein weiterer Unterschied ist das Vorhandensein eines Kiefergelenks zwischen Squa-mosum (Schädelknochen) und Surangulare (Unterkieferknochen) bei Probainognathus, während bei den Morganucodontiden das Squa-mosum mit dem Dentale ein Gelenk bildet. Die Tritheledontiden besitzen zahlreiche Gemeinsamkeiten mit den Morganucodon-tiden. So fehlt bei ihnen wie bei den Säugetieren ein präfrontaler und postorbitaler Knochen. Das ovale Fenster (Ort der Schallwellenübertragung von den Mittelohrknöchelchen auf das Innenohr) liegt ganz innerhalb der knöchernen Ohrkapsel (Perioticum) wie bei den Säugetieren. Im Gegensatz dazu ist bei allen anderen Cynodontiern diese Öffnung ganz oder teilweise durch zwei andere Schädelknochen (Basi-sphenoid und Basiocciput) umgeben. Völlig unähnlich den Morganucodontiden ist jedoch die ganz eigene Struktur der Backenzähne der Tritheledontiden (Abb. 8).
Die Tritylodontiden sind eine dritte, sehr fortschrittliche Cynodontiergruppe. Sie ist als letzte Therapsidenfamilie ausgestorben (Abb. 3). Das gut bekannte Körperstamm- und Extremitätenskelett (Abb. 9) weist Ähnlichkeiten mit den Morganucodontiden auf, deren postcraniales Skelett eine Kombination von Säugetiergrundstrukturen und primitiven cynodontierähnlichen Merkmalen darstellt. Der Schädel verfügt über Charakteristika der Morganucodontiden wie das Fehlen eines von der Augenhöhle abgegrenzten Schläfenfensters (Abb. 10) und eine ähnliche Struktur der Seitenwand der Hirnschale. Es gibt aber auch große Unterschiede zu den Morganucodon-tiden, insbesondere in der Struktur der Backenzähne. Auf den Backenzähnen sind die Höcker in Längsreihen angeordnet (Abb. 11), wodurch die Zähne beim Zahnschluss wie eine Feile wirken. Zusammenfassend lässt sich feststellen, dass zwischen den fortschrittlichsten säugerähnlichen Reptilien und den frühen Säugetieren erhebliche morphologische Unterschiede bestehen. Dieser Befund ist allgemein anerkannt. Einige Paläontologen bezweifeln sogar eine Abstammung der Morganucodontiden von den Cynodontiern überhaupt. Als ein Argument für diese Sicht wird der Verlauf einer der Hauptvenen des Kopfes (Vena capita lateralis) angeführt. Diese Vene verläuft bei den Morganucodontiden unterhalb und bei den Cynodontiern oberhalb des seitlichen Vorsprungs des Felsenbeinknochens. Der eine Zustand sei vom anderen nicht ableitbar. Werden die Morganucodontiden von primitiveren Therapsiden als den Cynodontiern hergeleitet, dann ist die zu überbrückende morphologische Kluft natürlich noch größer als bei der Ableitung von den Cynodontiern.
Als wichtigstes Argument für die Evolution der Säugetiere aus den Reptilien wird der für viele Autoren überzeugende Nachweis der Entstehung des Säugetiermittelohres mit drei Gehörknöchelchen aus reptilischen Vorfahren mit nur einem Gehörknöchelchen angeführt. Da bei diesem Strukturwandel phylogenetische Vorhersagen aufgrund embryologischer Befunde (Reichert-Gauppsche Theorie, Æ V.10.5.2) im nachhinein durch paläontologische Befunde scheinbar bestätigt worden sind, gilt die Phylogenese des Säugetiermittelohres als ein herausragender Beleg für die Evolutionslehre. Der Grundgedanke der Reichert-Gauppschen Theorie (V.11.6.2 im Lehrbuch) ist die Homologisierung (V.10.1) des Hammer-Amboss-Gelenks des Mittelohrs der Säugetiere mit dem „primären“ reptilischen Kiefergelenk zwischen Quadratum (Schädelknochen) und Artikulare (Unterkieferknochen) (Abb. 11.21 im Lehrbuch; vgl. Abb. 4A). Da das „primäre“ Kiefergelenk der Reptilien ins Mittelohr der Säugetiere gewandert sei, habe sich als Ersatz ein neues „sekundäres“ Kiefergelenk zwischen dem Squamosum (Schädelknochen) und dem Dentale (Unterkieferknochen) entwickelt. Eine fossile Dokumentation dieses Strukturwandels muss als Hauptschritte die Entstehung eines „sekundären“ Squamosum-Dental-Gelenks, die Ablösung des Artikulare vom Unterkiefer und des Quadratum vom Schädel und ihre Verlagerung ins Mittelohr mit Etablierung als Hammer und Amboss dokumentieren. Außerdem muss sich das Angulare aus dem Unterkiefer lösen, um als tympanischer Knochen, der das Trommelfell hält, mit dem Schädel zu verwachsen. Es darf aber nicht vergessen werden, dass zahlreiche weitere knöcherne, knorplige, bindegewebige, muskuläre und nervale Veränderungen parallel mit diesem Umwandlungsprozess einhergehen müssen. Bevor auf die vorhandenen fossilen Dokumente eingegangen wird, sei zunächst festgestellt, dass die Rekonstruktion des Hörapparates der fortschrittlichen Cynodontier (und anderer Therapsiden) und früher Säugetiere mit einer cynodontierähnlichen Unterkieferstruktur wie die der Morganucodontiden strittig ist. Im Mittelpunkt dieser Diskussion steht die Lokalisation des Trommelfells. Von einigen Forschern wird es hinter dem Quadratum wie bei den heutigen Reptilien rekonstruiert. Andere meinen, dass es im Unterkiefer hinter dem Dentale gelegen hat. Als dritte Möglichkeit wird postuliert, dass beide Anordnungen des Trommelfells gleichzeitig ausgeprägt waren. Auf diese Diskussion kann im Rahmen dieses Lehrbuches natürlich nicht eingegangen werden. Nur soviel sei bemerkt, dass die Art der Trommelfell-rekonstruktion die postulierten Evolutionsschritte zum Säugermittelohr beeinflusst. Davon unberührt bleiben aber die o.g. notwendigen evolutionären Hauptschritte. Auf der Basis des oben skizzierten postulierten Umbauprozesses stellt sich zunächst die Frage nach der Struktur des Unterkiefers und des Kiefergelenks bei den relevanten fossilen Formen. In zeitlicher Abfolge zeigen die Unterkiefer der Cynodontier und einiger früher Säugetiere eine zunehmende Vergrößerung des Dentale und eine Reduktion der nichtdentalen Unterkieferknochen, wobei der Kontakt zwischen dem Dentale und den postdentalen Knochen loser wird (Abb. 12). Das als Schädelknochen an der Gelenkbildung mit dem Unterkiefer (Artikulare) beteiligte Quadratum nimmt in dieser Reihe ebenfalls an Größe und in der Festigkeit des Kontaktes mit dem Schädel ab.
Bei bestimmten Cynodontiern gibt es neben dem „primären“ Kiefergelenk (Quadratum-Artikulare) einen zweiten Kontakt zwischen Schädel und Unterkiefer, das Squamoso-Surangular-Gelenk. Einige Paläontologen vermuten, dass das Squamoso-Surangular-Gelenk ein Zwischenstadium auf dem Wege zum Squamoso-Dental-Gelenk war. Dies ist aber eine völlig spekulative Deutung. Mindestens ebenso plausibel kann das Squamoso-Surangular-Gelenk als einmalige, bei den heute lebenden Formen nicht vorkommende Struktur mit einer bestimmten Funktion ohne verwandtschaftliche Beziehung zum Squamoso-Dental-Gelenk verstanden werden. Es diente möglicherweise als Hindernis gegen eine Gelenkverlagerung nach hinten. Ähnliches bewirkt ein knöcherner Vorsprung bei „höheren“ Säugetieren. Bei einigen sehr fortschrittlichen Cynodontiern soll das Dentale einen Kontakt mit dem Squamosum gehabt haben. Damit sei der Entwicklungsbeginn des säugetiertypischen Kiefergelenkes gegeben. Spätere Untersuchungen konnten diesen Befund nicht bestätigen. Er bleibt somit bei den Therapsiden zumindest umstritten. Auch könnte der Kontakt zwischen dem Squamosum und dem Dentale bei den Tritheledontiden aufgrund von Strukturunterschieden nicht plausibel als homolog dem Squamoso-Dental-Gelenk der Säugetiere gedeutet werden. Allerdings besitzen frühe Säugetiere wie z.B. die Morganucodontiden neben dem „pri-mären“ Quadrato-Artikular-Gelenk ein „sekundäres“ Squamoso-Dental-Gelenk (Abb. 4). Nur dieser Sachverhalt liegt der Behauptung von Evolutionstheoretikern zugrunde, nach der die Transformation des Mittelohrs der Säuger aus dem Mittelohr und Unterkiefer der Reptilien fossil belegt sei. Interessanterweise konnte die Existenz von zwei Kiefergelenken auch bei heute lebenden Vögeln nachgewiesen werden. Für den angenom-menen viel komplizierteren Prozess der Umwandlung von Quadratum, Artikulare und Angulare zu Amboss, Hammer und dem tympanischen Knochen mit entsprechender Verlagerung vom Unterkiefer zum Schädel bzw. ins Mittelohr gibt es keine fossilen Belege. In diesem Zusammenhang werden Unterkieferknochen der beiden Säugetiere Amphitherium (Familie Amphitheriidae) und Peramus (Familie Peramuridae) aus dem Jura angeführt, die eine erste Stufe in der Loslösung von Artikulare und Angulare repräsentieren sollen. Es muss im Rahmen dieses Lehrbuches genügen, festzustellen, dass diese Deutung sehr spekulativ ist. Außerdem wäre eine ausreichende fossile Dokumentation der Säugetiermittel-ohr-evolution damit auch nicht gegeben. Ein im Jahr 2005 veröffentlicher Fossilfund eines Kloakentiers erzwingt neuerdings die schon zuvor diskutierte Schlussfolgerung, dass die säugertypischen Gehörknöchelchen konvergent entstanden sind. Die an sich schon unwahrscheinliche Umbildung von Kiefergelenkknochen wäre demnach mindestens zweimal unabhängig erfolgt – ein evolutionstheoretisch sehr unplausibles Szenario.
Die heute lebende Klasse der Säugetiere (Mammalia) wird in zwei große Gruppen unterteilt. Alle eierlegenden Säugetiere gehören zu den Atheria (auch Prototheria genannt) und alle nichteierlegenden Säugetiere zu den Theria.
Die Monotremata (Kloakentiere) sind die einzige rezente Ordnung der Atheria. Sie unterscheiden sich ganz wesentlich von allen anderen lebenden Säugetieren und sind auf Australien, Tasmanien und Neuguinea beschränkt. Sie umfassen nur zwei Familien, das Schnabeltier (Ornithorhynchus) und die Schnabeligel (Tachy-glossus und Zaglossus). Die Theria werden in zwei Großgruppen Marsupialia (Beuteltiere) und Plazentalia („Mutterkuchentiere“) unterteilt. Die Marsupialia verdanken ihren volkstümlichen Namen der Tatsache, dass die im winzigen unreifen Zustand geborenen Jungen in einem Beutel auf der Bauchseite der Weibchen für eine bestimmte Zeit beherbergt und genährt werden. Ein ganz charakteristischer Vertreter dieser Gruppe ist das Känguruh. Der Name der zweiten Theria-Großgruppe, der Plazentalia, leitet sich von der Tatsache her, dass bei ihnen eine leistungsfähige Ernährungsverbindung zwischen Mutter und Embryo, die Plazenta (Mutterkuchen) ausgebildet ist. Infolgedessen erreicht das Junge ein viel fortgeschritteneres Stadium bis zur Geburt als bei den Beuteltieren. Die Plazentalia sind unter den heute lebenden Säugetieren die verbreitetste und formenreichste Gruppe. Es gibt zwei Merkmale, in denen sich die heutigen Theria von den Atheria unterscheiden und die an Fossilien erkannt werden können. Zum einen ist es der Aufbau der Seitenwand des Hirnschädels und zum anderen die Höcker-anordnung auf den Kronen der Backenzähne. Bei den Atheria wird ein großer Teil der Seitenwand des Hirnschädels von der knöchernen Ohrkapsel (Perioticum), bei den Theria statt dessen von einem anderen Knochenelement, dem Alisphenoid, gebildet. Die Theria besitzen Backenzähne mit einer dreieckförmigen Anordnung der Haupthöcker. Die Monotremata als einzige heute vorkommende nichttherische Säugetierordnung sind allerdings im erwachsenen Alter zahnlos und besitzen statt eines Gebisses einen Hornschnabel.
AtherialinieDie schon besprochenen Morganucodontiden besitzen eine seitliche Hirnschale, die überwiegend durch die Ohrkapsel gebildet wird. Da außerdem die Höcker der Backenzähne nicht in einem Dreieck, sondern linear angeordnet sind, wird diese frühe Säugetierfamilie als Ahn einer unbekannten Gruppe, aus der sich die Atheria entwickelt hätten, angesehen. Aufgrund neuer Funde wurde dann eine Abstammung der Monotremata von Theriasäugern (s.u.) favorisiert. Aber auch diese Hypothese ist durch ein neu entdecktes Fossil sehr fragwürdig geworden. Evolutionäre Abstammungsvorstellungen der Monotremata werden durch den paläontologischen Befund nicht gestützt. TherialinieAus der Zeit der Trias bis zur frühen Kreide, in der die wesentlichen Schritte der Theriaevolution erfolgt sein sollen, gibt es nur wenig Material vom Schädel und Körperstamm- und Extremitätenskelett. Die relevanten Fossilfunde bestehen überwiegend aus Zähnen und Unterkieferknochen. Die Vorstellung der Evolution der Theria basiert deshalb größtenteils auf der Phylogenie der Zähne, insbesondere der taxonomisch besonders interessanten Molaren. Die „höheren“ Säugetiere besitzen einen sogenannten tribosphenischen Molarentyp (Abb. 13). Solch ein Molar besitzt eine dreieckige Anordnung der Haupthöcker, wobei diese Dreikante an den unteren und oberen Molaren umgekehrt gegenüberstehen und so ineinander greifen können. Dem unteren Dreikant (Trigonid) ist hinten jeweils noch ein niedriger Anhang (Talonid) angegliedert, der den Innenhöcker (Protoconus) des nächstfolgenden oberen Molaren als Gegenüber aufnimmt. Unter den Molaren vom tribosphenischen Typ gibt es aber große Strukturunterschiede. Evolutionsbiologen haben nun versucht, ältere fossil bekannte Molaren auf der Grund-lage ihrer Kronenstruktur und des sich daraus ableitenden Facettenmusters in eine phylogenetische Reihe einzuordnen, die zu jüngeren fossilen Molaren mit einer tribosphenischen Grundstruktur führt. Kuehneotherium (Familie Tinodontidae, Abb. 3) ist eine sehr frühe Säugetiergattung, die obere und untere Backenzähne mit drei Höckern in dreieckförmiger Anordnung besitzt und deshalb als frühester Ahne der Theria gedeutet wird. Es muss aber an dieser Stelle festgestellt werden, dass es bereits im Jura den Atheria zugeordnete Säugetiergruppen (Docodonta, Multituberculata) gibt, die hochkomplexe Okklusionsverhältnisse aufweisen, wie sie in ähnlicher Weise erst wieder bei Marsupialia und Plazentalia aus dem Tertiär angetroffen werden. Von Kuehneotherium ausgehend markieren nach evolutionärer Vorstellung Amphitherium und Peramus wichtige Schritte auf dem Entwicklungsweg zum tribosphenischen Molaren, der sehr früh von Aegialodon (Familie Aegialodontidae) repräsentiert wird. Amphitherium aus dem Mitteljura besitzt gegenüber -Kuehneotherium eine zusätzliche Grundscherfläche und einen etwas verlängerten Anhang (Talonid). Bei Peramus aus dem Oberjura ist der Anhang gegenüber Amphitherium besser entwickelt. Aegialodon ist nur von einem einzelnen inkompletten Zahn aus der frühen Kreide bekannt und besitzt gegenüber Peramus eine zusätzliche Abnutzungsfläche auf dem etwas größeren Anhang. Diese Abnutzungsfläche weist auf einen Höcker an der Innenseite (Protoconus) des gegenüberliegenden Zahnes wie bei den Theria hin.
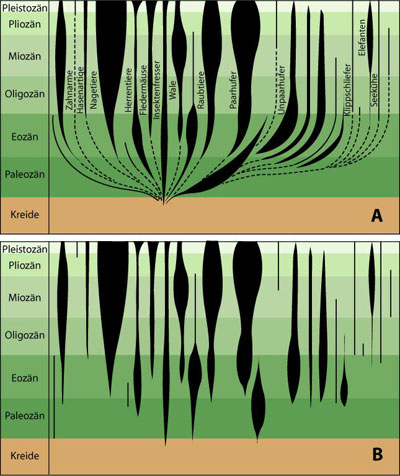
Grundsätzlich muss kritisch angefragt werden, ob selbst solch „kleine“ Strukturveränderungen in der vermuteten Evolutionsfolge des tribosphenischen Molaren wie die plötzliche Entstehung eines neuen Höckers mit zusätzlicher Scherfläche durch plausible Evolutionsmechanismen erklärt werden können. Von dieser grundlegenden Frage aber abgesehen, sehen auch Evolutionstheoretiker in der Reihe von Kuehneotherium bis Aegialodon keine reale phylogenetische Folge, sondern nur Vertreter typischer Evolutionsstufen. Der Grund dafür sind hochspezialisierte Merkmale bei jedem dieser Molaren, die sie als echte Ahnen der Theria ausschließen. Außerdem muss noch einmal darauf hingewiesen werden, dass von der wichtigsten Phase der angenommenen Theriaevolution wegen zu geringen Fossilmaterials vorwiegend nur die Molaren diskutiert werden können. Mit weniger Fossilmaterial sind im allgemeinen aber auch leichter stimmige Übergangsreihen konstruierbar als mit vielen Knochenüberresten, da bei weniger bekannten Merkmalen die Wahrscheinlichkeit von Merkmalskonflikten auch geringer ist (siehe Zahn- und Schädelmorphologie der fortschrittlichen Cynodontier und frühen Säugetiere). Neuere Fossilfunde legen mittlerweile nahe, dass sich tribosphenische Molaren zweimal völlig getrennt voneinander entwickelt hätten (STOKSTAD 2001). Dieser Backenzahntyp sei in der südlichen Hemisphäre unabhängig bei fossilen Verwandten der Monotremata, einer sehr alten Säugergruppe, aufgetaucht. Die fraglos tribosphenischen Molaren wurden Ende der 1990er Jahre in Australien und Madagaskar gefunden; sie gehören zu Ausktribosphenos und Ambondro. Zudem wurde Ambondro in einer mitteljurassischen Schicht gefunden, so dass der tribosphenische Molarentyp nicht nur in einer anderen Hemisphäre, sondern auch in einer anderen Zeit auftaucht, nämlich 10er von Millionen Jahren bevor die ersten Formen im Norden fossil auftreten. Es sind noch weitere Fossilfunde notwendig, um die Stichhaltigkeit einer getrennten Entstehung der beschriebenen Backenzahn-Typen bei den Säugern zu untermauern. Außer den säugerähnlichen Reptilien stehen also auch die verschiedenen Gruppen der frühen Säugetiere des Mesozoikums (Erdmittelzeit) durch deutliche morphologische Lücken getrennt nebeneinander. Ein plausibler Nachweis realer phylogenetischer Beziehungen existiert nicht. Dieser Befund gilt auch für die Fossilüberlieferungen sämtlicher 30 Säugetierordnungen der Erdneuzeit, die im Alttertiär (frühe Erdneuzeit) bzw. in der Oberkreide (späte Erdmittelzeit) getrennt voneinander auftreten (Abb. 14). Vielfältige frühe SäugetierfaunaBis Anfang der achtziger Jahre konnte man die Säugetierfossilien den Atheria und Theria bzw. Marsupialia und Plazentalia ziemlich eindeutig zuordnen. Neuere Funde zeigen, dass die frühe Säugetierfauna der Erdmittelzeit weit vielfältiger war als bis dahin angenommen und eine allein an den heute lebenden Formen orientierte Taxonomie als Einteilungsschema nicht mehr aufrecht erhalten werden kann. Außerdem zeigen diese Formen auch einige sehr fortschrittliche Merkmale zu einem sehr frühen Zeitpunkt. Solche Organismen stellen in einem evolutionären Kontext große Probleme dar, da sie bereits am Beginn der Säugerevolution Endprodukte dieser Entwicklung repräsentieren. Andererseits sind bei diesen Säugetieren auch ganz ursprüngliche und einmalige Merkmale nachweisbar. Es handelt sich teilweise um ausgeprägte Merkmalsmosaikformen. In der frühen Zeit der Säugetiere (Erdmittelzeit) tritt also eine größere Vielfalt an „gröberen“ Bauplänen auf als später (Erdneuzeit; dort gibt es allerdings eine deutlich größere Zahl von Familien als früher). Im folgenden werden fünf eindrucksvolle Beispiele genannt:
Dinnetherium (Familie Amphilestidae) aus dem Unterjura besitzt einerseits Zähne mit längsausgerichteten Höckern ähnlich den Morganucodontiden. Die Abnutzungsflächen auf den Backenzähnen weisen aber darauf hin, dass während des Zahnschlusses eine Verlagerung des Unterkiefers nach innen kombiniert mit einer Rotationsbewegung stattfand. Diese Bewegungen repräsentieren einen hohen Entwicklungsstand. Am Unterkiefer ist neben einem pseudangulären Fortsatz ein Knochenvorsprung in der angulären Region in Fortsetzung des Massetermuskels vorhanden (Abb. 15). Diese Unterkieferanatomie weist im Zusammenhang mit dem Zahnabnutzungsmuster auf einen oberflächlichen Massetermuskel mit ähnlicher Anordnung und Funktion wie bei den modernen Säugetieren hin. Mit Dinnetherium existiert also ein modernes Säugetiermerkmal in der frühen Phase der vermuteten Säuge-tier-evolution. Simpsonodon ist eine Gattung aus der Familie der Docodontidae aus dem Mitteljura. Simpsonodon besitzt den modernen Theria völlig vergleichbare Molaren, mit einem wichtigen Unterschied: Der Anhang (Talonid) ist am unteren Molar nicht hinten, sondern vorn lokalisiert. Sinoconodon (Familie Sinoconodontidae, Abb. 3, 5), ein anderes Säugetier aus dem Unterjura, besitzt einerseits fortschrittliche säugerähnliche Merkmale mit einem stark entwickelten Squamoso-Dental-Gelenk; der Condylus dentale ist groß und kugelig geformt und die gegenüberliegende Gelenkpfanne im Squamosum tief eingekerbt. Das Dentale ist groß und robust und die akzessorischen Unterkieferknochen kleiner als bei den anderen frühen Säugetieren. Andererseits besitzt Sinoconodon sehr primitive Merkmale vergleichbar mit dem nur mäßig entwickelten Cynodontier Thrinaxodon (Familie Thrinaxodontidae). So sind die Backenzähne nicht in Prämolaren und Molaren differenziert. Die Zähne kamen während des Kieferschlusses wahrscheinlich nicht in Kontakt. Das Zahnersatzmuster ähnelt dem von Cynodontiern. Andererseits ist aus der Kreide ein Krokodil mit ungewöhnlichen, fortschrittlichen Zähnen bekannt. Normalerweise besitzen die Krokodile relativ gleich aussehende kegelförmige Zähne. Dieses einmalige fossile Krokodil verfügt aber über komplexe säugetierähnliche Zähne. Die Haramiyiden (Abb. 3, 5) repräsentieren als sehr frühe Säugetiergruppe (Obertrias-Mitteljura) ebenfalls ein ausgeprägtes Merkmalsmosaik. Primitiv sind die großen postdentalen Unterkieferknochen. Die Zähne der Haramiyiden sind aber hochspezialisiert. Durch die Kieferschlussbewegungen wurde ein Bohr- und Brecheffekt erzielt, der sich vom Scher- oder Mahlmechanismus der anderen frühen Säugetiere völlig unterschied. Gobiconodon als fünftes Beispiel ist ein letzter Überlebender der Familie der Amphilestidae aus der Unterkreide. Dieses Säugetier ist wegen eines außergewöhnlichen Merkmals sehr interessant: Gobiconodon besitzt Zähne, die von ihrer Struktur und Funktion her Molaren sind, aber ersetzt werden. Dieser Befund widerspricht allen Beobachtungen an Säugetieren, nach denen Molaren keine vorausgehenden Zähne besitzen. Ein weiteres unerwartetes Merkmalsmosaik bietet ein fast vollständiges mausgroßes Skelett eines Säugetieres aus Liaoning in China von der Grenze Jura/Kreide (JI et al. 1999). Der Fund aus der Gruppe der Triconodonten mit dem Artnamen Jeholodens jenkinsi entspricht nicht unbedingt evolutionstheoretischen Erwartungen. Das postkranielle Skelett (unterhalb des Schädels) weist ein Merkmalsmosaik auf: Beckengürtel und Hinterextremität werden als primitiv eingestuft; der Schultergürtel dagegen als „sehr abgeleitet“, dem von fortschrittlichen Theriern gleichend (zu denen die heutigen Säugetierordnungen gehören). Für einen Fund dieses Alters ist dies im evolutionären Kontext unerwartet, denn Formen an der Basis der mutmaßlichen Evolution sollten eher durchgehend Primitivmerkmale aufweisen. JI et al. (1999) gehen daher davon aus, dass die fortschrittlichen Merkmale konvergent entstanden sind und damit keinen Hinweis auf einen Abstammungszusammenhang mit den modernen Säugetiergruppen geben. Sie äußern die Vermutung, dass nicht nur im Schädelbereich (wo das schon länger bekannt ist), sondern auch im Bereich des postkraniellen Skeletts mit häufigen Konvergenzen zu rechnen sei.
In jüngster Zeit wurde ein Säugerfossil gefunden, das an heutige termitenfressende Gürteltiere und Erdferkel erinnert (LUO & WIBLE 2005). Der älteste bekannte Termitenfresser Fruitafossor windscheffeli stammt aus dem Oberjura (der Zeit, aus der man den berühmten Urvogel Archaeopteryx kennt). Merkmale des Gebisses sowie die Grabbeine weisen bemerkenswerte Ähnlichkeiten mit den heutigen Termitenfressern auf. Da andere Merkmale gegen eine nähere Verwandtschaft sprechen, muss eine unabhängige Entstehung der speziellen Merkmale angenommen werden (Konvergenz). Fruitafossor ist ein weiteres Beispiel eines ursprünglichen Säugetiers mit sehr spezialisierten Merkmalen. Es besitzt zudem auch einzigartige Merkmale im Unterkieferbereich. Als letztes Fossil in dieser Reihe sei schließlich ein biberähnlicher Fund der Gattung Castorocauda aus dem Mitteljura genannt (JI et al. 2006). MARTIN (2006) bemerkt, dass dieses Fossil ein weiteres Puzzlestück in einer Reihe von Funden darstelle, die nicht wie erwartet primitiv und unspezialisiert seien. Besonders auffällig ist der abgeflachte Schwanz, der mit kleinen Hornschuppen bedeckt ist; die Hinterfüße besitzen Schwimmhäute, und die Bezahnung ähnelt fischfressenden Robben. MARTIN (2006) schließt seinen Kommentar wir folgt: „We stand at the threshold of a dramatic change in the picture of mammalian evolutionary history, and many chapters of it will soon need rewriting.“ Zeitliches ProblemBei dem Versuch, die Säugetiere von säugerähnlichen Reptilien plausibel abzuleiten, gibt es außer den genannten großen morphologischen Schwierigkeiten auch ein zeitliches Problem. Gegenwärtig werden am häufigsten die beiden Cynodontierfamilien Tritylodontidae und Thritheledontidae als nächste Verwandte (Schwesterntaxon) der Säugetiere eingestuft. Die Tritheledontiden werden stratigraphisch etwas älter als die Tritylodontiden eingeordnet. Das derzeit älteste bekannte Säugetier Adelobasileus (repräsentiert durch eine Hirnschale), wurde aber in Schichten geborgen, die nach herkömmlicher geologischer Zeitrechnung ca. 10 Millionen Jahre älter sind als die, aus denen die ältesten Tritheledontiden-Fossilien stammen (Æ Abb. 13.22). Man hofft dieses zeitliche Problem durch Funde von Tritylodontiden- und Tritheledontiden-Fossilien aus älteren Schichten zu lösen, was natürlich für die Zukunft nicht auszuschließen ist.
1. Das paläontologische Hauptargument für einen Übergang von Reptilien zu Säugetieren ist die Tatsache, dass von primitiven über fortschrittliche säugerähnliche Reptilien zu den frühen Säugetieren hin eine Formenreihe aufgestellt werden kann, die mit zunehmend jüngerem geologischen Alter auch eine größere Säugetierähnlichkeit aufweist. 2. Dabei dient die Unterkieferstruktur als Paradebeispiel für die Bestätigung der Evolutionslehre durch paläontologische Befunde. In zeitlicher Abfolge zeigen die Unterkiefer der Cyno-dontier und einiger früher Säugetiere eine zunehmende Vergrößerung des Dentale und eine Reduktion der nichtdentalen Unterkieferknochen, wobei der Kontakt zwischen dem Dentale und den kleineren postdentalen Knochen loser wird. Das als Schädelknochen an der Gelenkbildung mit dem Unterkiefer (Artikulare) beteiligte Quadratum nimmt in dieser Reihe ebenfalls an Größe und in der Festigkeit des Kontakts am Schädel ab. In der weiteren Entwicklung soll das „primäre“ reptilische Kiefergelenk durch ein „sekundäres“ Säugetierkiefergelenk zwischen dem Squamosum (Schädel) und dem Dentale (Unterkiefer) ersetzt worden sein. Mit der Befreiung von der Kiefergelenkfunktion sollen Quadratum und Artikulare in das Hammer-Amboss-Gelenk des Säugetiermittelohrs transformiert worden sein. Gleichzeitig sei das Angulare vom Unterkiefer zum Schädel gewandert, um dort die knöcherne Kapsel des Ohres (tympanischer Knochen) zu bilden. 3. Frühe Säugetiere, die neben einem reptilischen noch ein zweites säugetierähnliches Kiefergelenk besitzen, passen in das Evolutionskonzept. Diese Konstruktion muss jedoch nicht in diesem Rahmen gedeutet werden, sondern kann ohne eine phylogenetische Relevanz einfach nur funktionelle Ursachen haben. Für diese Deutung spricht das Auftreten von zwei Kiefergelenken bei einigen fortschrittlichen säugerähnlichen Reptilien und bei heute lebenden Vögeln. 4. Für den komplizierten Umwandlungsprozess von Quadratum, Artikulare und Angulare zu Säugetiermittelohrknochen gibt es keine fossile Dokumentation. Die wesentlichen Schritte der Evolution des Mittelohres der Säugetiere sind damit paläontologisch nicht nachgewiesen. 5. Die frühen Säugetiere und ihre vermuteten Ahnen unter den säugerähnlichen Reptilien besitzen viele Ähnlichkeiten. Beim Vergleich der Gesamtmorphologie zeigen sich aber auch große morphologische Unterschiede. So besitzen die am häufigsten als nächste Verwandte der frühen Säugetiere eingestuften Cynodontierfamilien Tritheledontidae und Tritylodontidae zwar säugerähnliche Merkmale im Schädelbereich, ihre Zähne sind aber denen der frühen Säugetiere völlig unähnlich. 6. Im Evolutionsmodell gibt es zusätzlich zu diesen morphologischen Schwierigkeiten ein zeitliches Problem. Die derzeit ältesten Funde der Tritheledontiden und Tritylodontiden werden geologisch jünger eingestuft als der derzeit älteste bekannte Säuger. Die Detailbetrachtung zeigt, dass in der Fossilüberlieferung plausible Bindeglieder beim Übergang vom Reptil zum Säuger fehlen. Auch in der vermuteten späteren Säugetierevolution lassen sich zwischen den zahlreichen Säugetiergruppen keine historisch-verwandtschaftlichen Beziehungen ableiten.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Studiengemeinschaft WORT und WISSEN e.V.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||