Zusammenfassung: In diesem Artikel wird erklärt, was in der Embryologie erforscht wird und weshalb dieses Forschungsgebiet in der Ursprungsfrage eine besondere Bedeutung genießt. In einem geschichtlichen Abriss wird besonders auf das Biogenetische Grundgesetz von Ernst Haeckel eingegangen und dessen Stellenwert für die Erforschung der Stammesgeschichte kritisch diskutiert. Inhalt
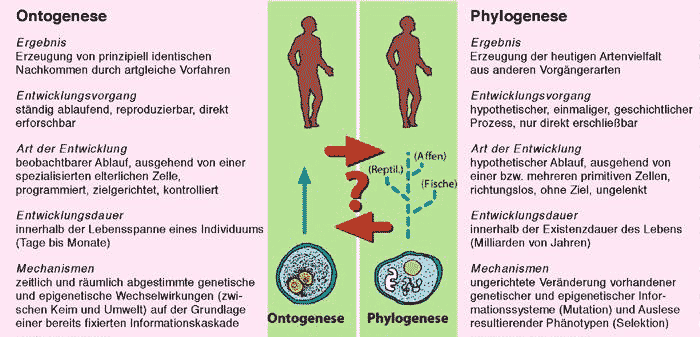
Eine wichtige Quelle für das Verständnis der Eigenschaften des Lebens liefern Untersuchungen, die den Bildungsweg mehrzelliger Organismen aus einer einzigen Zelle verfolgen. Im Gegensatz zur Phylogenese, bei der eine vermutete, nicht reproduzierbare Entwicklung von Arten auseinander im Verlauf langer Zeiträume Gegenstand der Forschung ist, handelt es sich bei der Ontogenese um den beobachtbaren Prozess der Bildung artgleicher Nachkommen (vgl. Abb. 1). Der dafür zuständige Wissenschaftszweig ist die Embryologie. Sie untersucht z. B. bei den Säugetieren den sich ständig wiederholenden Vorgang der Bildung von Organismen aus einer äußerlich undifferenziert erscheinenden, befruchteten Eizelle.
Aus praktischen Gründen werden zwei Bereiche der Embryologie unterschieden: die deskriptive (beschreibende) Embryologie und die kausale bzw. experimentelle Embryologie. Bei der ersten Forschungsrichtung geht es um die rein formelle, plastische Beschreibung der Entwicklungsvorgänge eines Organismus. Außerdem werden Vergleiche von Entwicklungsabläufen unterschiedlicher Arten auf struktureller und auf molekulargenetischer Ebene durchgeführt. Die kausale Embryologie bemüht sich um die Klärung von Ursachengefügen der dabei ablaufenden Bildungs-, Wachstums- und Differenzierungsprozesse. Ein Verständnis der Entwicklung eines Individuums (Ontogenese) und dessen Analyse im Vergleich mit den Vorgängen bei anderen Individuen ist nur aus der Zusammenschau der Ergebnisse beider Disziplinen möglich. Die Embryologie spielt auch für die Frage nach der Geschichte der Lebewesen eine bedeutsame Rolle. Kann der Entwicklungsweg einzelner Organismen als ein Abbild ihrer vermuteten Stammesgeschichte gelten? Sind vorübergehend auftretende Strukturen (z.B. die Pharyngealbögen beim Menschen) Relikte (= Überreste) von Merkmalen phylogenetischer (= stammesgeschichtlicher) Ahnen (hier: Kiemen der Fische)? Lässt sich eine ursächliche Verknüpfung von Stammesgeschichte und Ontogenese nachweisen? Welche Bedeutung zur Stammbaumrekonstruktion besitzen vergleichende embryonale Studien? Stellen genetische Entwicklungs- und Kontrollgene (Homeobox-Gene, vgl. vgl. III.6.3.9 im Buch), embryonale Bildungsmechanismen und -prinzipien gleichzeitig auch einen zentralen Angriffspunkt für evolutionäre Mechanismen des Artenwandels dar? Das sind einige Fragen, auf die in diesem Artikel und den beiden weiteren Artikeln zur Biogenetischen Grundregel eingegangen wird (vgl. Biogenetisches Grundgesetz – Aktuell und Biogenetisches Grundgesetz – Beispiele).
Ontogenese bezeichnet bei Tieren die durch die Verschmelzung von mütterlicher Ei- und väterlicher Samenzelle (Befruchtung) eingeleitete Individualentwicklung eines Organismus bis zu seiner Geburt. Zum Teil wird dieser Begriff auch erweitert angewendet auf die Keimes- und Jugendentwicklung (bis zur Geschlechtsreife) oder auf das gesamte Leben eines Organismus bis zu seinem Tod. Die Ontogenese des Menschen im Mutterleib wird in drei Phasen unterteilt. Die ersten zwei Wochen nach der Befruchtung werden Blastogenese genannt. In dieser Periode der menschlichen Frühentwicklung geschieht der Transport des menschlichen Keims durch den Eileiter und seine Einnistung in die Gebärmutter. Dabei bildet sich nach vielen Zellteilungen die kugelähnliche Blastozyste (Abb. 2). An einem Pol zeigt diese eine Zellverdichtung, aus welcher u.a. das Fruchtwasserbläschen (Amnionhöhle), die embryonale Plazenta und die Keimscheibe hervorgehen. Die Keimscheibe repräsentiert die erste Anlage des embryonalen Körpers des Menschen.
Der zweite Abschnitt, die Embryonalperiode, dauert von der 3. bis zur 8. Schwangerschaftswoche. In dieser Phase werden alle Organe angelegt und die „Körperform“ gestaltet. Der Mensch wird jetzt Embryo genannt (Abb. 3). Bedeutsam ist in dieser Phase auch die Bildung des Mutterkuchens (Plazenta), welcher im Verlauf der Entwicklung lebensnotwendige Funktionen (Ernährung, Atmung, Hormonbildung usw.) garantiert. Den letzten und längsten Abschnitt der Ontogenese von der 9. bis zur 40. Schwangerschaftswoche bezeichnet man als Fetalperiode. Sie ist durch das Wachstum und die funktionelle Reifung der einzelnen Organanlagen und Körperteile des Fetus gekennzeichnet und wird mit der Geburt abgeschlossen Entscheidend für den normalen Verlauf der Ontogenese sind vielfältige, zeitlich und örtlich exakt aufeinander bezogene Wechselbeziehungen auf genetischer, zellulärer, geweblicher und organischer Ebene. Diese sind innerhalb des kindlichen bzw. mütterlichen Organismus sowie zwischen ihnen nachweisbar. Eine zentrale Rolle für die Form- und Funktionsentwicklung besitzt das genetische Material des Kindes, welches väterliche und mütterliche Erbinformationen zu gleichen Anteilen enthält. Aber die 46 Chromosomen stellen nicht die alleinigen Informationsquellen des regulativen Einflusses auf die Gestaltungsvorgänge dar. Eiweißverbindungen im Zellkörper (Plasma) und komplizierte Oberflächenmoleküle der Zellmembranen können Signale mechanischer oder biochemischer Natur des Kindes oder der Mutter speichern, aufnehmen und abgeben. Diese Moleküle fungieren zum einen als Sender, zum anderen aber auch als Empfänger (Rezeptoren). Die zeitliche Abfolge aller Regulationsprozesse und der resultierenden einzelnen Entwicklungsschritte während der Ontogenese sind genau aufeinander abgestimmt. Jedes einzelne Glied in der ontogenetischen Kette ist zugleich Resultat vorangegangener Prozesse und Grundlage für andere Strukturen und Funktionen. Trotz des reichen Bestandes an Detailwissen zur menschlichen Ontogenese und von Modellorganismen wie der Fruchtfliege, des Fadenwurms, des Zebrafisches, des Hühnchens oder der Maus sind viele Fragen noch ohne Antworten geblieben. Insbesondere ist die Bildung der morphologisch dreidimensionalen, funktionell ausgereiften Lebensform in wesentlichen Punkten noch unverstanden.
Bis zur Mitte des 18. Jahrhunderts verband man mit dem Begriff „Evolution" eine besondere Vorstellung zur Ontogenese der Organismen. Deren frühe Entwicklung wurde als eine Art Heranwachsen oder „Auswickeln“ (lat. evolvere) eines bereits im Samen komplett ausgebildeten Individuums betrachtet (Einschachtelungs- bzw. Präformationslehre). Gegen diese Sichtweise wandte sich Caspar Friedrich Wolff (1733-1794). In seiner herausragenden Arbeit „Theoria Generationis“ von 1759 demonstrierte er anhand der Entwicklung des Hühnchens im Ei, dass Entwicklung kein bloßer Wachstumsprozess eines bereits ausgestalteten Organismus ist, sondern von einer Aufeinanderfolge vielfältiger Strukturneu- und -umbildungen bestimmt wird. Seine Epigeneselehre (Epigenese: Entstehung völlig neuer Strukturen während der Embryonalentwicklung) markiert den Beginn der modernen Embryologie. Sie konnte sich jedoch erst 50 Jahre später gegen die Präformationslehre durchsetzen. Die vergleichende Embryologie wurde Ende des 18. Jahrhunderts durch die Stufenleiterlehre und die Typologie bestimmt. Beide Anschauungen waren mit naturphilosophischen Vorstellungen eng verknüpft (Oken 1779-1851, Goethe 1749-1832, Schelling 1775-1854). Nach der Stufenleiterlehre (Abb. 4) sollten alle Pflanzen und Tiere in lückenlosen linearen Reihen (Stufenleitern) entsprechend ihres Komplexitätsgrades einzuordnen sein. Dabei wurde eine Übereinstimmung systematischer Reihen der Embryologie zu denen der Paläontologie (= Fossilforschung) angestrebt. Die Entwicklung des Menschen im Mutterleib, der an der Spitze aller Stufenleitern stand, war nach Johann Friedrich Meckel d.J. (1781-1833) ein schrittweises Durchlaufen aller unter ihm stehenden Organismen, beginnend bei einfachen Formen (Weichtiere, Fische) und zu den komplizierteren (Landtiere, Affen) aufsteigend. Andere Gelehrte vertraten die Idee des Typus. Bedeutendster Vertreter dieser Anschauung auf dem Gebiet der vergleichenden Anatomie war Georges Cuvier (1769-1832). Alle Organismen lassen sich danach auf bestimmte morphologische Grundmuster reduzieren. Für die jeweiligen systematischen Ebenen (Stamm, Klasse, Ordnung, Familie) suchte man nach einem entsprechend abgestuften abstrakten Bauplantyp. Der berühmte Embryologe Carl Ernst von Baer (1792-1876) wandte sich 1828 entschieden gegen die Grundgedanken der Stufenleiterlehre. Im Gesetz der Embryonenähnlichkeit postulierte er, dass in der Entwicklung eines Organismus zunächst die allgemeinen Bauplanmerkmale seines Stammes erscheinen und erst am Ende die speziellen, arttypischen Strukturen gebildet werden. Deshalb wären Embryonen der Wirbeltiere in den ersten Entwicklungsphasen nicht voneinander zu unterscheiden. Dies mit einer Art Rekapitulation, als Wiederholung von Bauplänen primitiver historischer Ahnen, im Sinne der Abstammungslehre gleichzusetzen, lag nie in den Absichten von Baers. Deutlich sprach er sich gegen die Deszendenzlehre (= Abstammungslehre) Darwins aus. Elemente des Stufenleiterdenken und Typologie finden sich später bei Ernst Haeckel vereinigt in einer phylogenetischen (= stammesgeschichtlichen) Lesart wieder.
Der deutsche Biologe Ernst Haeckel (1834-1919) zählt zu den wirkungsvollsten – aber auch umstrittensten – Forscherpersönlichkeiten des 19. und 20. Jahrhunderts. Sein herausragendes Engagement für die Darwinsche Deszendenztheorie (= Abstammungslehre) schuf die Basis für ihren rasanten Siegeszug in Deutschland. Haeckel bemühte sich nicht nur ehrgeizig um eine Neuinterpretation des biologischen Wissens seiner Zeit, sondern verwies sofort auf die sich daraus ergebenden weltanschaulichen Konsequenzen. Seine kompromisslos vertretene monistische Grundposition, die allein den Gesetzen der Natur Erklärungsanspruch auf alle Fragen des Menschen einräumte, steigerte er bis ins Extrem. Mit fanatischem Eifer verlangte Haeckel, alle naturwissenschaftlichen Fakten im Sinne der Abstammungslehre zu interpretieren. Ontogenetische Detailkenntnisse über zahlreiche Wirbeltiergruppen waren zu seiner Zeit kaum vorhanden. Dies überbrückte er wiederholt durch Präsentationen höchst spekulativer Schemazeichnungen, eigenen willkürlichen Modellierungen und überzogenen Verallgemeinerungen (vgl. Abb. 5 mit Abb. 6), auch wenn manche Tatsachen seinen Vorstellungen zur Phylogenese widersprachen. Dem Fälschungsvorwurf ausgesetzt schreibt Haeckel in Bezug auf die vielfach angezweifelten Abbildungen von Embryonen selbst (Berliner Volkszeitung vom 29. 12. 1908): „'Die gefälschten Embryonenbilder.' Um dem ganzen wüsten Streite kurzerhand ein Ende zu machen, will ich nur gleich mit dem reumütigen Geständnis beginnen, daß ein kleiner Teil meiner zahlreichen Embryonenbilder (vielleicht 6 oder 8 vom Hundert) wirklich (im Sinne von Dr. Braß) 'gefälscht' sind – alle jene nämlich, bei denen das vorliegende Beobachtungsmaterial so unvollständig oder ungenügend ist, daß man bei Herstellung einer zusammenhängenden Entwicklungskette gezwungen wird, die Lücken durch Hypothesen auszufüllen, und durch vergleichende Synthese die fehlenden Glieder zu rekonstruieren.“
Im Kontext dieses Zitats verteidigte Haeckel seine „Fälschungen", indem er die Methode des Schematisierens bei vielen seiner Kollegen ebenso nachweisen konnte und damit seine Abbildungen formell legitimiert sah. Diese Entschuldigung entspricht aber nicht der ganzen Wahrheit. Zeitgenössische Anatomen wie v. Baer, Rathke (1793-1860), Reichert (1811-1883), Kölliker (1817-1905) oder His (1831-1904) haben im 19. Jahrhundert in ihren Arbeiten zur Embryologie der Wirbeltiere oder des Menschen Tatsachen geliefert, die Haeckel einfach ignorierte bzw. nicht berücksichtigte, um seine Sicht der Dinge widerspruchsloser darstellen zu können. Auch wenn Haeckel wiederholt darauf hinwies, dass ein Teil seiner Zeichnungen hypothetisch waren und deshalb nicht der Realität entsprechen konnten, werteten und werten andere Fachkollegen diese Form der Darstellung als Fälschung oder Betrug (z.B. Rütimeyer 1868, His 1875, Bischoff 1876, Richardson 1997, Gould 2002). Der Fälschungsvorwurf zielte im Kern nie auf methodische, sondern auf inhaltliche Missgriffe. Mit den kritisierten Schemata bildete Haeckel nicht den Konsens bekannten Wissens ab, sondern entstellte Bekanntes und deklarierte Unbekanntes als bekannt mit dem Ziel, das Biogenetische Grundgesetz zu beweisen. Ungeachtet des nicht unerheblichen z.T. auch unsachlichen und polemischen Widerstandes vieler Berufskollegen sowie religiös motivierter Gegner (z.B. Kepler-Bund) blieb sein Einfluss auf Lehrbücher, die Morphologie und die Embryologie wirksam bis in die heutige Zeit hinein. Das lag im wesentlichen daran, dass in den vielen Diskussionen um Haeckel seine Thesen oder „Gesetze“ sowie seine Arbeitsweise und Person als Wissenschaftler mit dem Siegeszug der Abstammungslehre gleichgesetzt wurde und dadurch eine besondere Form von Legitimation erfuhr.
Haeckel sah in den Befunden der Individualentwicklung die „bedeutendsten Lichtträger für das Verständnis unserer Stammesgeschichte“. Zwischen Ontogenese und Phylogenese konstruierte er durch das Biogenetische Grundgesetz „naturgesetzliche“ Zusammenhänge. Allerdings findet sich keine klare, durchgängig gleichlautende Definition des Biogenetischen Grundgesetzes bei Haeckel. In seinen zahlreichen Werken existiert eine Vielzahl von Formulierungen mit zusätzlichen Ergänzungen und Einschränkungen. Zum ersten Mal stellte Haeckel 1866 seine Ansichten in der „Generellen Morphologie der Organismen" (2. Band, S. 300f.) unter der Überschrift „Thesen vom Causalnexus der biontischen und der phyletischen Entwicklung“ vor. 1872 erhob er diese Thesen zum Biogenetischen Grundgesetz (Die Kalkschwämme – Calcispongae, Berlin 1872). Theoretisches Fundament für das Biogenetische Grundgesetz war die über jeden Zweifel erhabene Überzeugung Haeckels von der Richtigkeit der Abstammungslehre. „Die Descendenz-Theorie ... bedarf zu ihrer vollen Gültigkeit keiner weiteren Beweise“ (1866, S. 294). Im folgenden werden die wichtigsten inhaltlichen Grundlinien von Haeckels Verständnis des Wechselverhältnisses zwischen Ontogenese und Phylogenese angeführt, unterteilt in deskriptive und kausale Aussagen.
1. Die Phylogenese ist die mechanische Ursache der Ontogenese. Haeckel wertete den Ablauf der Ontogenese eines Organismus als unmittelbares Resultat seiner Stammesgeschichte. Für beide Entwicklungsprozesse machte er dieselben Mechanismen („Vererbung und Anpassung“) verantwortlich. 2. Das Auftreten von Rekapitulation in der Ontogenese wird durch die Mechanismen der Phylogenese erzeugt. In der Ontogenese eines höherentwickelten Organismus sollen vorübergehend morphologische Strukturen auftreten, die adulten (= ausgereiften, erwachsenen) Merkmalen stammesgeschichtlicher Vorfahren entsprechen. Diese Wiederholungen nannte Haeckel Rekapitulationen. Er ging davon aus, dass neue, im Verlauf der Phylogenese erworbene Merkmale meist am Ende der Ontogenese sichtbar werden. Stammesgeschichtlich ältere Merkmale würden daraufhin mehr und mehr in ein früheres Stadium der Embryonalentwicklung verdrängt. (Später nannte man diesen hypothetischen Mechanismus „ terminale Addition“.) In der Ontogenese eines höheren Wirbeltieres fänden sich deshalb Merkmale primitiver Vorfahren am Anfang und die der unmittelbaren Vorgänger am Ende seiner Individualentwicklung (vgl. Abb. 7).
3. Der kausale Mechanismus: Vererbung erworbener Eigenschaften. Haeckel meinte, dass Vererbung auf der direkten Weitergabe erworbener Eigenschaften – von den Elterntieren an die Nachkommen – beruht. (Ähnlich dachten auch Lamarck und Darwin.) Besondere Eiweißstoffe innerhalb der Zelle, Plastidule, hätten die Fähigkeit der Reproduktion und verfügten über eine Art Gedächtnis (in Form periodischer Molekularbewegungen). Dieses würde durch die Umweltbedingungen des Organismus ständig verändert und abgewandelt. Die so geprägten Eiweißstoffe, die über die Keimzellen auf die nächste Generation übertragen werden sollten, ließen dann bei den Nachkommen entsprechende umweltadaptierte Merkmale entstehen.
1. Die Ontogenese des Individuums repräsentiert verkürzt die wichtigsten Formveränderungen der stammesgeschichtlichen Vorfahren (Adultformen) der eigenen Art. Danach repräsentieren einzelne embryonale Stadien jeweils ausgewachsene (adulte) Formen entsprechender Vorfahren. Im Zusammenspiel mit der Paläontologie und der vergleichenden Anatomie lässt sich somit der phylogenetische (= stammesgeschichtliche) Werdegang des jeweiligen Organismus exakt rekonstruieren. 2. Es existiert kein vollständig identisches Abbild der Phylogenese durch die Ontogenese. „Die vollständige und getreue Wiederholung der phyletischen durch die biontische Entwicklung wird verwischt und abgekürzt . . . und abgeändert“ (Haeckel 1866, S. 300). Die Embryonalentwicklung kann nicht in jedem Detail als völlig identisches Abbild der Phylogenese gelten, schränkte Haeckel von Anfang an ein. Es müsse deshalb unterschieden werden zwischen Palingenesen, die frühere stammesgeschichtliche Stadien während der Individualentwicklung wiederholen (Rekapitulation), und den Caenogenesen, ontogenetisch notwendige, aber stammesgeschichtlich nicht verwertbare Neubildungen. Palingenetische Merkmalsbildungen sind die bereits erwähnten Rekapitulationen. Dazu zählte Haeckel z.B. beim Menschen die sogenannten Kiemenbögen und die Schwanzanlage. Die Entstehung des Dottersackes, der Allantois, der Plazenta, der Eihüllen (z.B. Amnion) und der Nabelschnur sind dagegen caenogenetisch zu bewertende Strukturen. Auch die Verlagerung des ursprünglichen Erscheinungsortes eines Merkmales (Heterotopien) und Verschiebungen in der zeitlichen Reihenfolge des Auftretens von Organen (Heterochronien) im Vergleich mit der zugrundegelegten Phylogenese desselben Organs berücksichtigte Haeckel. 3. Die Ontogenie repräsentiert erst allgemeine und später spezielle Merkmale der Art. Als wichtige Ergänzung zum Biogenetischen Grundgesetz bezeichnete Haeckel das „Gesetz des ontogenetischen Zusammenhangs systematisch verwandter Thierformen“. In Anlehnung an die typologischen Vorstellungen von v. Baer (1828) im Gesetz der Embryonenähnlichkeit formulierte er: „Der Mensch ist demnach in den frühen Entwicklungsstufen nicht von den Embryonen der Vögel und Reptilien zu unterscheiden. Wenn auf noch frühere Stadien der Entwicklung zurückgegangen wird, wären keine Unterscheidungen zwischen den Embryonen dieser höheren Wirbelthiere und denjenigen der niederen, der Amphibien und Fische, aufzufinden“ (Anthropogenie 1877, S. 295). In den Abbildungen Haeckels erscheint dieser Zusammenhang besonders nachhaltig in der bereits angesprochenen „schematisierten“ Art und Weise (vgl. Abb. 5). Schlussfolgerungen. Ausgehend vom Paradigma „Evolution“ suchte Haeckel mit dem Biogenetischen Grundgesetz „den engen, ursächlichen Zusammenhang ...“ (Welträthsel 1903, S. 36) zwischen Phylogenese und Ontogenese darzustellen. Damit wollte er gleichzeitig einen methodischen Leitfaden für die Stammbaumrekonstruktion festschreiben. In den Grundgedanken des Biogenetischen Grundgesetzes und den dazugehörenden Ergänzungen und Ausnahmeregelungen finden sich frühere naturphilosophische Anschauungen (Stufenleiter, Typologie) in einer phylogenetischen Interpretation wieder. Haeckel schuf ein deskriptives und kausales Argumentationssystem, das flexibel, faszinierend und plausibel erschien. Aufgrund unverkennbarer Zirkularitäten, falscher Grundannahmen, überzogener Schlussfolgerungen (s.u.) und zweckorientierter Darstellungsformen musste sich der Jenaer Zoologe lebenslang mit wissenschaftlicher und persönlicher Kritik auseinandersetzen. Durch das Biogenetische Grundgesetz verlieh er der vergleichenden Embryologie jedoch einen enormen Forschungsstimulus. Gemeinsam mit Carl Gegenbaur (1826-1903) etablierte er in Deutschland das phylogenetische Denken als einzig akzeptablen Weg für die Bewertung des Datenmaterials aus der vergleichenden Anatomie und Embryologie.
Vom Zeitpunkt der Formulierung des Biogenetischen Grundgesetzes an gab es heftige Kontroversen über dessen inhaltlichen und methodischen Wert für die Biologie. Nur wenige Forscher übernahmen und verbreiteten seine Vorstellungen komplett und unverändert (Krause, Overzier, Bölsche). Die Mehrzahl der Biologen berücksichtigte nur einzelne Grundgedanken bzw. reduzierten ihre Bedeutung. So wurde z.B. nicht mehr von einem Gesetz, sondern von der biogenetischen Grundregel gesprochen (Weismann, Naef, Remane, de Beer, S. J. Gould). Eine nicht minder bedeutende Anzahl von Wissenschaftlern lehnte das Biogenetische Grundgesetz konsequent in jeder Hinsicht ab (His, Rütimeyer, Blechschmidt, D. S. Peters, Richardson). Problematisch erwies sich vor allem der methodische Ausgangspunkt Haeckels, die Abstammungslehre als bewiesene Tatsache zu deklarieren. Dies musste zwangsläufig zu Zirkelschlüssen bei der Interpretation ontogenetischer Daten führen. Im Einzelnen können folgende Kritikpunkte zu den kausalen Aussagen des Biogenetischen Grundgesetzes formuliert werden:
1. Die Ontogenese ist unabhängig von Phylogenese verstehbar. Träfe der kausale Ansatz Haeckels zu, wäre ein Verständnis des Formwandels, der Differenzierungs- und Wachstumsprozesse während der Ontogenese nur durch das Erforschen der Phylogenese möglich. Dagegen argumentierte bereits His (1868), der die Prozesse der Bildung von Organstrukturen unter Berücksichtigung mechanischer Gesetzmäßigkeiten zu erklären suchte. Die Gleichsetzung der Evolutionsmechanismen mit den für die Ontogenese verantwortlichen Vorgängen verwarf auch Kölliker (1884) mit dem Hinweis, „daß die Darwinsche Anpassungslehre nicht genügt, um die Umbildung der niederen Formen zu erklären, und ... daß die Entwicklungsgesetze der Organismen noch gänzlich unbekannt“ sind. Blechschmidt (1961 und 1973) konnte durch umfangreiche Detailstudien zeigen, dass für die Ontogenese des Menschen die Voraussetzung „Phylogenese“ nicht benötigt wird, um verstehbar zu sein. Die arttypische Formbildung im Verlauf der Ontogenese des Menschen führte er auf differenziert agierende Stoffwechselfelder zurück (siehe Anhang). Im weiteren dokumentiert die in den letzten Jahrzehnten erfolgte Integration molekulargenetischer und experimenteller Erkenntnisse zur Beschreibung entwicklungsbiologischer Vorgänge eindrucksvoll die Unabhängigkeit des Verständnisses der Ontogenese von phylogenetischen Modellvorstellungen. Während für Haeckel die Ontogenese nur aus der Phylogenese erklärbar schien, favorisiert die Evolutionsforschung heute den umgekehrten methodischen Weg: Vom Verständnis der Ontogenese her sucht sie nach den kausalen Ursachen des phylogenetischen Wandels. 2. Ontogenetische und phylogenetische Bildungsprozesse unterliegen nicht den gleichen Mechanismen. Weismann (1834-1914), selbst ein überzeugter Anhänger der Abstammungslehre, demonstrierte die Unhaltbarkeit der Vererbung erworbener Eigenschaften in einem berühmt gewordenen Experiment. Er schnitt vielen Generationen von Mäusen die Schwänze ab, ihre Nachkommen wurden jedoch niemals schwanzlos geboren. Die Wiederentdeckung der von Gregor Mendel 1865 formulierten Vererbungsgesetze gegen Ende des 19. Jahrhunderts offenbarte deutlich die Falschheit der Haeckelschen Vorstellungen. Ebenso spricht das gegenwärtige Wissen bezüglich der genetischen Grundlagen von Vererbungsprozessen gegen die dem Biogenetischen Grundgesetz zugrundeliegenden Annahmen. Die experimentelle Embryologie entdeckte, aufbauend auf der von Roux (1850-1924) am Anfang des 20. Jh. begründeten Entwicklungsmechanik, weitere vielfältige Zusammenhänge der raumzeitlichen Steuerung der Entwicklungsvorgänge sowie der Induktion und Differenzierung im Zusammenspiel von Genotyp und Phänotyp. Die Ontogenese ist die gezielte Realisierung der im genetischen (DNS) und epigenetischen Raum (Zellkern, Plasma, Zellmembran) zum Zeitpunkt der Befruchtung vorhandenen Informationen zur Erzeugung eines artgleichen Individuums. Die Erforschung embryonaler Bildungsvorgänge erfährt gegenwärtig durch das wachsende Wissen über die sogenannten Entwicklungssteuerungsgene (z.B. Homeobox-Gene, vgl. III.6.3.9 im Buch) einen enormen Aufschwung. Die dabei erkannten Zusammenhänge raum-zeitlicher Regulationskaskaden zeigen keine Gemeinsamkeiten mit den Vorstellungen zur Entstehung evolutionärer Neubildungen.
1. Die Gleichsetzung des Verlaufs der Ontogenese eines Organismus mit seiner Phylogenese ist nicht möglich. Haeckels Behauptung, dass die Gesamtheit der Formveränderungen eines Organismus während der Ontogenese denen seiner eigenen Stammesgeschichte entspricht, konnte durch vergleichende ontogenetische Studien nicht bestätigt werden. Bereits gegen Ende des 19. Jahrhunderts zeigten Keibel (1896) und Mehnert (1896), dass solche Parallelisierungen bei weitem nicht den Tatsachen entsprechen. Verschiebungen des zeitlichen Erscheinens und des örtlichen Auftretens (Heterochronien, Heterotopien) vieler Organe in der Ontogenese gegenüber ihrem vermuteten phylogenetischen Entstehen sind keine seltenen Ausnahmen, sondern die Regel. Auch der Vergleich von Ontogenesen stammesgeschichtlich eng verwandter Organismen (z.B. bei den Amniota (= Tiere, die ein Amnion = Embryonalhülle besitzen)) zeigt ausgeprägte Verschiebungen, zum Beispiel beim Erscheinen der Anlagen für das Herz oder der Augen (Jeffery et al. 2002). Deren Wertung im Sinne einer Rekapitulation würde zu absurden Ergebnissen führen. Diese Unterschiede werden mittlerweile mit neuen Methoden der vergleichenden Forschung („event-pairing") für die Stammbaumrekonstruktion analysiert.
Aufgrund dieser Situation beschränkte man sich weitestgehend darauf, >Rekapitulationen nur noch auf Organebene nachzuweisen. „Wenn es Rekapitulationen gibt, so betreffen sie keinesfalls vollständige Ahnenformen (wie primär angenommen wurde), sondern allenfalls einzelne Organe“ (Sudhaus & Rehfeld 1992, S. 159).
2. In der Ontogenese eines Individuums treten keine Adultmerkmale (= ausgereifte Merkmale des erwachsenen Organismus) stammesgeschichtlicher Vorfahren auf. Die morphologischen und physiologischen Besonderheiten embryonaler und fetaler Strukturen lassen eine pauschale homologe Gleichsetzung mit adulten Merkmalen rezenter und hypothetischer stammesgeschichtlicher Vorfahren nicht zu. Zum Beispiel finden sich bei der embryonalen Ausbildung des vorderen Darmabschnittes der Reptilien und Säugetiere niemals die funktionstüchtigen Kiemen mit Kiemenspalten oder Kiemenblättchen der angenommenen Ahnen. Dieser Körperteil wird phylogenetisch aus dem Kiemendarm der Fische und Amphibien abgeleitet. Naef (1917) begründete deshalb in seinem „Gesetz der konservativen Vorstadien“, dass nur der Vergleich von embryonalen Organanlagen bei Individuen, die derselben phylogenetischen Linie angehören, für die Stammbaumforschung sinnvolle Schlussfolgerungen erlauben. 3. Objektive Maßstäbe zur Unterscheidung von Palingenesen und Caenogenesen fehlen. Eine zweifelsfreie Kennzeichnung ontogenetischer Strukturen als palingenetisch oder caenogenetisch kann nicht aus ihnen selbst heraus erschlossen werden. (Palingenese = Wiederholung hypothetischer Vorfahrenstadien; Caenogenese = Neubildung, die nicht als stammesgeschichtliche Wiederholung gedeutet werden kann; s. o.) Denn „die Entscheidung, ob in einer bestimmten Entwicklungsphase eine Caenogenese oder aber eine Palingenese vorliegt, kann erst dann exakt getroffen werden, wenn die Phylogenese (= Stammesgeschichte, Abstammungsabfolge) bekannt ist“ (Siewing 1987, S. 271). Diese Voraussetzung ist allerdings nicht gegeben, da bis heute der vollständige stammesgeschichtliche Wandel von keiner einzigen Art bekannt ist. „... und mit der objektiven Unsicherheit auf diesem Gebiete wächst die Kraft der subjektiven Überzeugung und damit die Heftigkeit der wissenschaftlichen Fehden“ (Spemann 1915, S. 75). Diese Einschätzung besitzt auch heute noch ihre volle Gültigkeit. Die Diskussion um die Herleitung der Gehörknöchelchen aus den Kieferknochen der Reptilien ist dafür exemplarisch (vgl. Biogenetisches Grundgesetz – Beispiele). 4. „Vom Allgemeinen zum Speziellen“ stellt nicht den Weg der Ontogenese dar. Beim Vergleich immer jüngerer, „primitiverer" ontogenetischer Entwicklungsstadien verschiedener Organismen, z. B. bei den Wirbeltieren, soll eine zunehmende Ähnlichkeit der Keime nachweisbar sein (o. g. Gesetz der Embryonenähnlichkeit nach C. E. v. Baer bzw. „Gesetz des ontogenetischen Zusammenhangs systematisch verwandter Thierformen" nach Haeckel). Die Ontogenese eines Individuums verläuft danach über Stufen, bei dem zunächst einfache bzw. allgemeine und erst später spezieller konstruierte Formen der Organe und Strukturen wie in der Phylogenese sichtbar werden. Auch diese These ist widerlegt worden. Die Ontogenese lässt sich nicht mit einer Art aufsteigenden Entwicklungsprozess vom Allgemeinen zum Speziellen gleichsetzen. So nimmt die menschliche Entwicklung ihren Ausgang mit einer höchst spezialisierten Zelle mit 46 Chromosomen und nicht auf einer dem Pantoffeltierchen gleichzustellenden Ebene. Analoges gilt auch für die anderen Erscheinungsformen (z.B. Blastozyste) des menschlichen Keims.
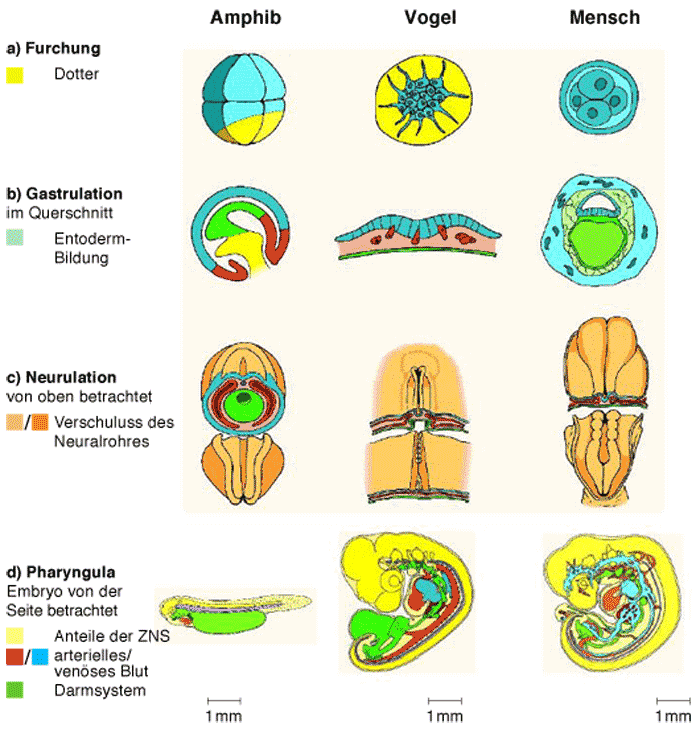
Außerdem kann bei den Wirbeltieren von einer zunehmenden Ähnlichkeit der Keime beim Vergleich immer jüngerer ontogenetischer Entwicklungsstadien keine Rede sein. Die Furchungstypen der befruchteten Eizelle, die Blastula bzw. Blastozystenbildung, die Gastrulation und die Mesodermbildung sowie die Neurulation zeigen bei Amphibien, Reptilien, Vögeln und Säugern deutliche Unterschiede (Abb. 8, Abb. 9 und Abb. 10). Unter Berücksichtigung dieser Erkenntnisse wurde dennoch an der Idee festgehalten, Stadien zu definieren, denen eine hohe phylogenetische Bedeutung zukommt. Die Entwürfe zur „Körpergrundgestalt“ (Seidel 1960, vgl. Abb. 9 unterste Reihe), eines phyletischen (Cohen 1993) oder des phylotypischen (Sander 1983) Stadiums oder des Phylotypus (Slack et al. 1993) und des Zootypus (Hall 1992) sind Ausdruck dieses Bemühens, beobachtbare Ähnlichkeiten zwischen den Embryonen in verschiedenen Entwicklungsphasen mit besonderer phylogenetischer bzw. palingenetischer Bedeutung zu belegen. Da jedoch ein methodisches Kriterium fehlt, um mit Sicherheit ein Stadium oder ein Merkmal gegenüber anderen als „universeller“ oder „konservierter“ zu kennzeichnen (s.o.), ist es unmöglich, sichere Homologisierungen festzulegen. Schließlich ist der Grad an Ähnlichkeiten in den einzelnen Entwicklungsphasen unterschiedlicher Wirbeltierarten entgegen dem „Sanduhrmodell“ (Abb. 11) in dem als „konservativ“ vermuteten Stadium nicht größer als in allen anderen. Breit angelegte vergleichende Studien führten zu dem Ergebnis, dass in keiner Phase der frühen embryonalen Entwicklung bei Wirbeltieren von einem phylogenetisch konservierten Stadium gesprochen werden kann (Richardson 1995, 1997). Dies steht nicht im Widerspruch zu der besonders günstigen Homologisierbarkeit einzelner Strukturen in der Embryonalperiode (zum Begriff „Homologie" und dessen Kritik vgl. V.10.1 im Buch). Im angesprochenen Stadium der Körpergrundgestalt zeigen die Embryonen der Fische, Amphibien, Reptilien, Vögel und Säugetiere gemeinsam die Anlage der Chorda dorsalis, der Visceralbögen, der Extremitätenknospen und der caudalen (= am Körperende gelegenen) Knospe. Aus makroskopischer Sicht mögen diese Keime verschiedener Wirbeltiere zu diesem Zeitpunkt nahezu deckungsgleich erscheinen. Die Gedanken von v. Baer und Haeckel sind bei den damals noch gering entwickelten optischen Möglichkeiten durchaus nachvollziehbar. Wendet man jedoch eine der Größe des zu untersuchenden Objektes (1-5 mm) angepasste Auflösung und Vergrößerung an, werden auch auf mikroskopischer Ebene des Embryonalstadiums erhebliche Unterschiede deutlich. Diese sind nach Richardson zum Teil ausgeprägter als bei den ausgewachsenen Tieren (Abb. 6), z.B. hinsichtlich ihrer Größe (bis zum 10 fachen), ihres gesamten Bauplanes (Zahl der Somiten, Pharyngealbögen bzw. Gliedmaßen) oder in der Anlage bzw. des Differenzierungsgrades einzelner Organe (z.B. des embryonalen Herzens).
Ernst Haeckel hat sein Biogenetisches Grundgesetz als wissenschaftlich begründete Darstellung und direkten methodischen Ansatz zur Aufklärung des als eng und ursächlich eingeschätzten Zusammenhanges zwischen der Phylogenese und Ontogenese proklamiert. Die Embryologie konnte weder die damit verknüpften kausalen noch dessen deskriptive Aussagen bestätigen. Das Biogenetische Grundgesetz in der Version Haeckels war bereits am Anfang des 20. Jahrhunderts wissenschaftlich widerlegt. Als eigenständige methodische Grundlage für die Phylogeneseforschung konnte es aufgrund der begrifflichen und inhaltlichen Unschärfe niemals dienen. Dem Namen nach überlebte es dennoch, wenn auch in stark eingeschränkter und modifizierter Form (z.B. als „biogenetische Grundregel“) in der vergleichenden Embryologie. Dabei beruft man sich vor allem auf einzelne deskriptive Inhalte des Biogenetischen Grundgesetzes, die auf Einzelorgane bezogen und im Rahmen der Homologieforschung (vgl. V.10.1 im Buch) entsprechend ausgewertet werden.
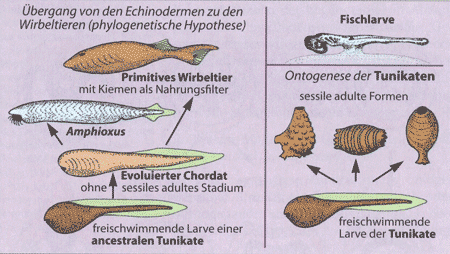
Die Methode der direkten Übertragung von Ontogenesestadien eines Organismus oder Stadien einzelner Organe auf deren phylogenetischen Werdegang wird gelegentlich weiterhin praktiziert (z. B. Ableitung der Chordatiere aus Tunikatenlarven; siehe Abb. 12). Als wichtige und von vielen auch als einzig mögliche Methode wird dagegen der indirekte Weg angesehen. Dabei wird zunächst akzeptiert, dass ein unmittelbares Ablesen der Stammesgeschichte anhand ontogenetischer Abläufe nicht möglich ist. Beim indirekten Weg werden ontogenetisch auftretende Merkmale mit embryonalen und adulten (= ausgereiften) Strukturen anderer Arten verglichen, um Homologien zu ermitteln (vgl. V.10.1 im Buch). Die auf diese Weise bestimmten Homologien können evolutionstheoretisch u. U. als Rekapitulationen gedeutet und damit als Hinweise auf stammesgeschichtliche Zusammenhänge verstanden werden. Beispiele für so ermittelte Rekapitulationen sind die embryonalen Zahnanlagen bei Bartenwalen, die Anlage des primären Kiefergelenkes bei Säugetieren, die Chorda dorsalis bei den Säugern, die Bildung von Krallen beim Schopfhuhn-Nestling (Hoatzin) oder die Änderung der Lobenlinien im Verlauf der Ontogenese bei Ammoniten. Die als Rekapitulation gedeuteten Entwicklungen gelten häufig als „Umwegentwicklungen“, da sie scheinbar „unnötig kompliziert“ verlaufen. Die embryonale Anlage der Visceral- oder Pharyngealbögen („Kiemenbögen“), der Chorda dorsalis (elastischer Stab aus Knorpelgewebe) oder des Ductus Botalli (Blutgefäß zwischen Lungen- und Körperschlagader) bei Säugetieren erscheint danach unökonomisch und historisch bedingt. Nur bestimmte Restfunktionen bzw. die Übernahme anderer, sekundärer Aufgaben im Organismusgefüge hätten sie im Verlauf der Evolution konserviert. Diese Erklärungen verlieren jedoch beim gegenwärtigen Stand der Forschung mehr und mehr an Boden. So dient z.B. die Chorda bei Säugetieren als Platzhalter und Induktor für die spätere Bildung der Wirbelsäule und des Neuralrohres. Die Zahnanlagen bei zahnlosen Säugetieren sind unverzichtbar für die formgerechte Kieferentwicklung. Der Nutzen vorübergehend in der Ontogenese auftretender Organe ist offensichtlich. „Im Rahmen ihrer Stoffwechselbewegungen haben alle Organe, Gewebe und Zellen Gestaltungsfunktionen. Funktionslose Organe oder Atavismen finden wir nicht. Schon die frühesten pränatalen Organsysteme funktionieren, und zwar gemäß den Eigenschaften, die sie in den jeweiligen Phasen ihrer Entwicklung haben“ (Blechschmidt 1996, S. 77). Dabei dürfen Organe nie isoliert betrachtet werden, um ihre funktionelle Bedeutung richtig einzuschätzen.
Jede normale Differenzierung vollzieht sich in biodynamischen Stoffwechselfeldern mit räumlich geordneten Stoffwechselbewegungen (siehe zum folgenden Abb. 13). Auflockerungs(Sog-)felder (a): Zonen, in denen während des Wachstums durch äußere Zugkräfte ein Sog entsteht. Durch Einsprossen von Epithelzellen in das Auflockerungsgebiet differenzieren sich hier regelmäßig Drüsen. Contusionsfelder (Stauchung, b): Frühe Skelettanlagen, in denen unter Druck von außen Zellverbände zusammengepresst werden. Auf diese Weise entstehen z.B. im Vorknorpel die charakteristischen, zunächst tellerförmigen jungen Knorpelzellen.Corrosionsfelder (Zerreißung, c): Wenn zwei Gewebe so eng aufeinandergepresst werden, dass zwischen ihnen kein gefäßführendes Gewebe Platz hat, erlischt die Nahrungszufuhr und die Zellen gehen zugrunde. Derartige Stoffwechselfelder sind Corrosionsfelder. In ihnen sterben die Zellen ab und eröffnen Kommunikationen flüssigkeitsgefüllter Hohlräume. Densationsfelder (Verdichtung, d): Die Entstehung des embryonalen Skeletts ist bestimmt durch Verlust an wässriger Grundsubstanz. Flüssigkeit diffundiert ab, der Zellverband wird dadurch verdichtet. Detraktionsfelder (Gleiten mit Reiben, e): Stoffwechselfelder, in denen Flüssigkeit durch Gleitbewegung von Zellen mit starker Reibung entlang von Gleitflächen schnell ausgepresst wird. Detraktionsfelder sind die Entstehungsgebiete von Knochen. Dilationsfelder (Dehnung, f): Zonen, in denen Zellen in bestimmten Hauptrichtungen auf Zug beansprucht und dabei gedehnt werden. So dilatierte Zellen entwickeln sich gesetzmäßig zu kontraktionsfähigen Muskelzellen. Distusionsfelder (Stemmen, g): Knorpelzellen können auf Grund ihrer Lage ihre Abbauprodukte nur schwer an die Umgebung abgeben. Durch die gestauten Abbauprodukte bekommen Knorpelzellen osmotische Eigenschaften – sie quellen und üben Stemmkörperfunktionen aus. Retensionsfelder (Straffung, h): Binnengewebe, das durch Querdruck beengt und durch einen senkrecht dazu verlaufenden Zug gestrafft wird, bekommt Zugfestigkeit und funktioniert damit als Halteapparat. Sehnen und Bänder entstehen in Retensionsfeldern.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Studiengemeinschaft WORT und WISSEN e.V.
|